Tadalafil entfaltet seine Wirkung über eine selektive Hemmung der PDE5, wodurch die Konzentration von cGMP im glatten Muskelgewebe stabil bleibt. Diese biochemische Modulation resultiert in einer langanhaltenden Relaxation der Gefäßwände. Der Wirkstoff wird nach oraler Einnahme effizient resorbiert, mit einer Bioverfügbarkeit von rund 80 %. Seine Halbwertszeit von bis zu 36 Stunden ist innerhalb dieser Substanzklasse außergewöhnlich. Abgebaut wird er in der Leber, hauptsächlich durch CYP3A4, mit anschließender biliärer Exkretion. Typische unerwünschte Wirkungen entstehen durch eine verstärkte Vasodilatation, etwa Kopfschmerzen oder Flush. Pharmakologisch wird cialis generika vor allem durch die verlängerte Wirkungsdauer charakterisiert.
Anti-age-adn.eu
Brief Communication Reconstitution of telomerase activity in normal human cells leads to elongation of telomeres and extended replicative life span Homayoun Vaziri and Samuel Benchimol Normal somatic cells have a finite life span [1] and lose
and pBabest2-AS retroviruses were transfected into the
telomeric DNA, present at the ends of chromosomes,
packaging cell line Phoenix-E and viral supernatants were
each time they divide as a function of age in vivo or in
harvested. The normal human diploid fibroblast strain BJ,
culture [2–4]. In contrast, many cancer cells and cell
previously transfected with the ecotropic virus receptor
lines established from tumours maintain their telomere
gene [18], was infected with viral supernatants (multiplic-
length by activation of an RNA–protein complex called
ity of infection = 4) at approximately 75–79 population
telomerase, an enzyme originally discovered in
doublings (PDs). These cells have approximately 10–15
Tetrahymena [5], that synthesizes telomeric repeats
PDs remaining before reaching senescence. Colonies
[6–8]. These findings have led to the formation of the
resistant to both G418 and hygromycin were selected,
‘telomere hypothesis’, which proposes that critical
isolated with cloning cylinders and expanded. BJ cells,
shortening of telomeric DNA due to the end-replication
like other fibroblasts, do not normally have telomerase
problem [9] is the signal for the initiation of cellular
activity [7], but they do express the RNA subunit (hTR) of
senescence [10,11]. In yeast, the EST2 gene product, the
the telomerase complex [15]. We reasoned, therefore, that
catalytic subunit of telomerase, is essential for telomere
BJ cells, which normally reach senescence after 87–90
maintenance in vivo [12–14]. The recent cloning of the
PDs, represented a suitable recipient cell strain in which
cDNA encoding the catalytic subunit of human
to express hTERT, reconstitute telomerase activity, and
telomerase (hTERT) [15,16] makes it possible to test the
test for elongated telomeres and extended life span. telomere hypothesis. In this study, we expressed hTERT in normal human diploid fibroblasts, which lack
The pBabe-infected and pBabest2-AS-infected BJ cells
telomerase activity, to determine whether telomerase
formed sparse and small colonies (Figure 1a,b). In contrast,
activity could be reconstituted leading to extension of
pBabest2-infected cells gave rise to many larger colonies
replicative life span. Our results show that retroviral-
(Figure 1c). Ten drug-resistant colonies, each consisting of
mediated expression of hTERT resulted in functional
approximately 250–350 cells, were isolated from each of
telomerase activity in normal aging human cells.
the infected cultures and expanded. In this secondary
Moreover, reconstitution of telomerase activity in vivo
plating assay, none of the 20 colonies derived from the
led to an increase in the length of telomeric DNA and to
pBabe-infected or pBabest2-AS-infected cultures was
extension of cellular life span. These findings provide
capable of dividing sufficiently to reach confluence even
direct evidence in support of the telomere hypothesis,
after 14–20 days. These cells appeared to have reached
indicating that telomere length is one factor that can
senescence after an estimated 87–90 PDs on the basis of
determine the replicative life span of human cells.
increased cell size, failure to divide and increased endoge-nous β-galactosidase activity (Figure 2a,b) [19]. In contrast,
Address: Ontario Cancer Institute and Department of Medical
9 of 10 colonies obtained from the pBabest2-infected
Biophysics, University of Toronto, 610 University Avenue, Toronto,Ontario, M5G 2M9, Canada.
Correspondence: Homayoun VaziriE-mail: vaziri@oci.utoronto.ca
Received: 9 January 1998 Revised: 26 January 1998 Accepted: 26 January 1998
Published: 16 February 1998 Current Biology 1998, 8:279–282 http://biomednet.com/elecref/0960982200800279
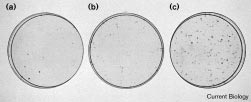
Colony-forming ability of BJ cells infected with retroviruses encoding the
Results and discussion
human telomerase catalytic subunit, hTERT, and controls. BJ cells
The cDNA encoding hTERT was subcloned in the retro-
infected with (a) pBabest2-AS, (b) pBabe or (c) pBabest2 were
viral vector pBabe under the control of the promoter
replated and selected in G418 and hygromycin. Cells were fixed and
present in the Moloney murine leukemia virus long
stained with methylene blue to detect colonies. The total number ofcolonies (containing ≥ 400 cells) in duplicate dishes was: pBabest2-AS,
terminal repeat [17] in both sense (pBabest2) and anti-
sense (pBabest2-AS) orientation. The pBabe, pBabest2
Current Biology, Vol 8 No 5
Endogenous β-galactosidase activity in BJ cells infected with retroviruses encoding the human telomerase catalytic subunit, hTERT, and controls. Cells were infected with (a) pBabest2-AS (assessed at PD ~87), (b) pBabe (assessed at PD ~87) and (c) pBabest2 (assessed at PD 117).
cultures reached confluence in the secondary plating assay.
and PD 123 (Figure 3). Extracts prepared from varying
To test for extended proliferative potential, single cells
numbers of cells indicated that the level of telomerase
were isolated by limiting dilution from the nine surviving
activity in the TIELF cells was comparable to that of the
clones and grown in a tertiary plating assay. These cells
established adenovirus5-transformed human cell line 293
have now been in continuous culture and have reached a
(Figure 3). Telomerase activity was sensitive to RNase and
minimum of 116 PDs (Table 1). The mean PD number for
the 10 pBabest2-infected clones was 28 PDs higher thanthe 20 control clones (p < 10–5 by student’s t-test). No evi-
Telomere length was measured using the terminal restric-
dence of β-galactosidase activity has been detected in
tion fragment (TRF) length assay as described previously
these dividing cultures (Figure 2c). To date, these cultures
[4]. Genomic DNA was extracted from BJ cells and from
show no sign of senescence and continue to divide. The
two independent TIELF clones at several PDs and the
cells show no sign, so far, of aneuploidy and, like normal
mean TRF length was determined. As expected, control
fibroblasts, they arrest their growth in response to low
BJ cells lose telomeric DNA with each PD at a rate of
serum and undergo contact inhibition at high density
–76 bp/PD (Figure 4a,b), similar to rates that we reported
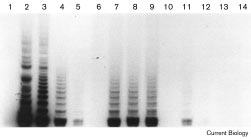
BJ cells infected with pBabest2 had reconstitutedtelomerase activity as detected by the telomeric repeatamplification protocol (TRAP) assay (Figure 3), or by theconventional primer extension assay (data not shown),whereas BJ cells infected with pBabe or pBabest2-AS hadno detectable activity (Figure 3). Reconstitution of telom-erase activity has also been observed in other telomerase-negative cells [20]. We refer to the pBabest2-infectedclones as TIELF cells (for telomerase-induced extendedlife span human fibroblasts). TIELF cells continuouslyexpressed telomerase activity over time at PD 111, PD 117
Reconstitution of telomerase activity in BJ human diploid fibroblasts. Extended life span of telomerase-positive clones.
TRAP assays were performed as described previously [24]. Lane 1,RNase-treated 293 cell extract serving as a negative control; lanes
2–5, 293 cell extracts corresponding to 105, 104, 103 and 102 cells,respectively; lane 6, RNase-treated TIELF cell extract; lanes 7–9, cell
extracts derived from 103 TIELF cells at PD 111, PD 117 and PD 123,respectively; lane 10, TIELF cell extract heated at 80°C for 3 min; lanes
11,12, cell extract corresponding to 100 and 50 TIELF cells,
respectively, at PD 123; lanes 13,14, cell extracts corresponding to105 BJ cells infected with pBabest2-AS and pBabe, respectively.
*PDs reached by pBabe and pBabest2-AS represent the maximum PD. Brief Communication
Analysis of TRFs in BJ and BJ-derived TIELFcells. Genomic DNA, purified by DNAzol
(BRL), was digested with HinfI and RsaI,
quantitated in triplicate by fluorometry and
1 µg DNA per lane was resolved on a 0.5%
described [4]. (a) Lanes 1–3, DNA size
markers; lanes 4,5, blank; lane 6, young BJ
cells at PD 29; lane 7, BJ cells at PD 51; lane
8, old BJ cells at PD 87; lanes 9–12, TIELF1
cells with increasing PDs as indicated; lanes
13,14, BJ cells infected with control (C)pBabest2-AS and pBabe, respectively, at PD
~50–55. (b) Quantitative analysis of telomeric DNA in normal BJ cells and the TIELF cells.
Normal BJ cells lost telomeric DNA at a rate
of –76 bp/PD, r = –0.98. Two TIELF cell
clones increased their telomere length at a
rate of +40 bp/PD, r = –0.96 and +94 bp/PD,
previously in these cells [21]. However, TIELF1 and
applications. Ectopic expression of telomerase in normal
TIELF2, two clones derived from pBabest2-infected BJ
cells may be successfully used in gene therapy to increase
cells, acquired very long telomeres rapidly in the initial
the life span of cells carrying the desired transgene.
expansion phase that continued to elongate with increas-
Similar approaches can also be used for treatment of
ing PD number; the approximate rates were +40 bp/PD
and +94 bp/PD in clones TIELF1 and TIELF2, respec-tively (Figure 4a,b). An increase in the size and intensityof the TRF signal from TIELF1 is evident from the
Materials and methods Cell cultureThe neonatal human fibroblast cell strain (BJ) attained a maximum lifespan of approximately 87–90 PDs under our conditions. Cells were
Our results provide direct evidence for the telomere
grown in α-minimal essential medium supplemented with 10% foetal
hypothesis. They show that forced expression of hTERT
bovine serum (FBS). Older cells were grown in medium supplemented
cDNA in normal human cells results in telomerase
with 15% FBS. Cells were split at a ratio of 1:4 or 1:8 at early passageor at a ratio of 1:2 in later passages. Phosphate-buffered saline con-
activity, elongation of telomere length and an extended
tained no calcium or magnesium. Plating efficiency for BJ cells was
life span. Thus, normal human cells can bypass the
> 90%. PD number was calculated by the count/split method or as
Hayflick limit and increase their replicative life span upon
PD = log(Nf/N0)/log2, where Nf is the final cell number and N0 is the
expression of telomerase activity. It is notable that the
length of telomeric DNA was not merely maintained but
increased in TIELF cells. After submission of this manu-
The retroviral constructs were packaged using the highly efficient and
script, similar results were reported by Bodnar et al. [22].
helper-free cell lines Phoenix-A and Phoenix-E (ATCC). Packaging
In contrast to their study, in which young/midlife cells
cells were transfected when approximately 80% confluent. Phoenix
were used to extend life span, our study used older cells
cells were incubated in 25 µM chloroquine 5 min prior to transfection
that had completed 80% of their life span and, hence,
with 10 µg retroviral plasmid DNA by the calcium phosphate tech-nique. At 72 h post-transfection, the virus-containing medium was col-
demonstrates that senescence can be prevented even in
lected and the virus titre determined using NIH3T3 cells. Titres of
old cells. Our results are reminiscent of studies with germ
2–4 × 106 transducing units per ml were obtained. BJ cells were
cells, which, unlike somatic cells, express telomerase
infected in the presence of polybrene (4 µg/ml) using viral super-
activity, have long TRFs (~15 kb) and show a net increase
natants at a multiplicity of infection = 4. Throughout this work, thetransfection efficiencies were monitored by a cytomegalovirus–
in the length of the telomeric DNA with age [23]. These
enhanced green fluorescent protein (CMV–EGFP) construct. BJ cells
results suggest that expression of telomerase in normal
were incubated for 20 h at 32°C in virus-containing medium. Fresh
human cells might be associated with processes related to
media was added and the cells were incubated for a further 24 h in
de-differentiation and could lead to generation of cells
virus-free media prior to trypsinization and plating in media supple-mented with G418 (400 µg/ml) and hygromycin (20–50 µg/ml). Drug-
with the stem cell property of indefinite self renewal.
resistant colonies were isolated with cloning rings approximately 2–3
TIELF cells could replace genetically unstable, estab-
weeks later. Throughout this work, BJ cells carrying pM5-Eco (the
lished cell lines currently used in a wide variety of
Current Biology, Vol 8 No 5 Acknowledgements
23. Allsopp RC, Vaziri H, Patterson C, Goldstein S, Younglai EV, Futcher
This work was supported by grants from the Medical Research Council of
AB, et al.: Telomere length predicts replicative capacity of human fibroblasts. Proc Natl Acad Sci USA 1992, 89:10114-10118.
Canada and the National Cancer Institute of Canada. We thank Greg Morin
24. Kim NW, Wu F: Advances in quantitation and characterization of
and Calvin Harley for the hTRT cDNA and Garry Nolan for providing the
telomerase activity by telomeric repeat amplification protocol
retroviral protocols used in this study. (TRAP). Nucleic Acids Res 1997, 25:2595-2597. References
1. Hayflick L, Moorhead P: The serial cultivation of human diploid strains. Exp Cell Res 1961, 25:585-621.
2. Harley CB, Futcher AB, Greider CW: Telomeres shorten during ageing of human fibroblasts. Nature 1990, 345:458-460.
3. Hastie ND, Dempster M, Dunlop MG, Thompson AM, Green DK,
Allshire RC: Telomere reduction in human colorectal carcinoma and with ageing. Nature 1990, 346:866-868.
4. Vaziri H, Schachter F, Uchida I, Wei L, Zhu X, Effros R, et al.: Loss of telomeric DNA during aging of normal and trisomy 21 human lymphocytes. Am J Hum Genet 1993, 52:661-667.
5. Greider CW, Blackburn EH: Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell 1985, 43:405-413.
6. Counter CM, Avilion AA, LeFeuvre CE, Stewart NG, Greider CW,
Harley CB, Bacchetti S: Telomere shortening associated with chromosome instability is arrested in immortal cells which express telomerase activity. EMBO J 1992, 11:1921-1929.
7. Kim NW, Piatyszek MA, Prowse KR, Harley CB, West MD, Ho PL, etal.: Specific association of human telomerase activity with immortal cells and cancer. Science 1994, 266:2011-2015.
8. Morin GB: The human telomere terminal transferase enzyme is a ribonucleoprotein that synthesizes TTAGGG repeats. Cell 1989, 59:521-529.
9. Olovnikov AM: A theory of marginotomy. Doklay Biochem 1971, 201:394-397.
10. Harley CB: Telomere loss: mitotic clock or genetic time bomb? Mutat Res 1991, 256:271-282.
11. Harley CB, Vaziri H, Counter CM, Allsopp RC: The telomere hypothesis of cellular aging. Exp Gerontol 1992, 27:375-382.
12. Lendvay TS, Morris DK, Sah J, Balas B, Lundblad V: Senescence mutants of Saccharomyces cerevisiae with a defect in telomere replication identify three additional EST genes. Genetics 1996, 144:1399-1412.
13. Counter CM, Meyerson M, Eaton EN, Weinberg RA: The catalytic subunit of yeast telomerase. Proc Natl Acad Sci USA 1997, 94:9202-9207.
14. Lingner J, Hughes TR, Shevchenko A, Mann M, Lundblad V, Cech TR:
Reverse transcriptase motifs in the catalytic subunit of telomerase. Science 1997, 276:561-567.
15. Nakamura TM, Morin GB, Chapman KB, Weinrich SL, Andrews WH,
Lingner J, et al.: Telomerase catalytic subunit homologs from fission yeast and human. Science 1997, 277:955-959.
16. Meyerson M, Counter CM, Eaton EN, Ellisen LW, Steiner P, Caddle
SD, et al.: hEST2, the putative human telomerase catalytic subunit gene, is up-regulated in tumor cells and during immortalization. Cell 1997, 90:785-795.
17. Morgenstern JP, Land H: Advanced mammalian gene transfer: high titre retroviral vectors with multiple drug selection markers and a complementary helper-free packaging cell line. Nucleic Acids Res 1990, 18:3587-3596.
18. Albritton LM, Tseng L, Scadden D, Cunningham JM: A putative murine ecotropic retrovirus receptor gene encodes a multiple membrane-spanning protein and confers susceptibility to virus infection. Cell 1992, 57:659-666.
19. Dimri GP, Lee X, Basile G, Acosta M, Scott G, Roskelley C, et al.: A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc Natl Acad Sci USA 1995, 92:9363-9367.
20. Weinrich SL, Pruzan R, Ma L, Ouellette M, Tesmer VM, Holt SE, et al.:
Reconstitution of human telomerase with the template RNA component hTR and the catalytic protein subunit hTRT. Nat Genet 1997, 17:498-502.
21. Vaziri H, West MD, Allsopp RC, Davison TS, Wu YS, Arrowsmith CH,
et al.: ATM-dependent telomere loss in aging human diploid fibroblasts and DNA damage lead to the posttranslational activation of p53 protein involving poly(ADP-ribose) polymerase. EMBO J 1997, 16:6018-6033.
22. Bodnar AG, Ouellette M, Frolkis M, Holt SE, Chiu C-P, Morin GB, etal.: Extension of life-span by introduction of telomerase into normal human cells. Science 1998, 279:349-352.
Information and prices are correct at the time of publication (July 2011), however may be subject to change. *P.O.A – Please phone 1300 552 512 for clarification of the fee. $33.15 (Medicare rebate available under certain circumstances) thromboembolism or First degree relative who has a prove defect of Antithrombin, protein C/S or APCR ADH $30.70 (Invoice from $31.15 (Invoi
 Brief Communication
Brief Communication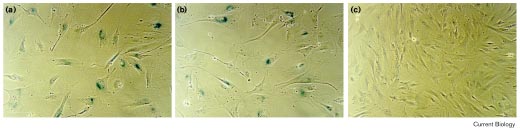
 Current Biology, Vol 8 No 5
Current Biology, Vol 8 No 5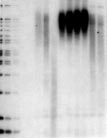 Brief Communication
Brief Communication