Tadalafil entfaltet seine Wirkung über eine selektive Hemmung der PDE5, wodurch die Konzentration von cGMP im glatten Muskelgewebe stabil bleibt. Diese biochemische Modulation resultiert in einer langanhaltenden Relaxation der Gefäßwände. Der Wirkstoff wird nach oraler Einnahme effizient resorbiert, mit einer Bioverfügbarkeit von rund 80 %. Seine Halbwertszeit von bis zu 36 Stunden ist innerhalb dieser Substanzklasse außergewöhnlich. Abgebaut wird er in der Leber, hauptsächlich durch CYP3A4, mit anschließender biliärer Exkretion. Typische unerwünschte Wirkungen entstehen durch eine verstärkte Vasodilatation, etwa Kopfschmerzen oder Flush. Pharmakologisch wird cialis generika vor allem durch die verlängerte Wirkungsdauer charakterisiert.
Pii: s0169-409x(97)00108-7
Advanced Drug Delivery Reviews 30 (1998) 73–83
Recent advances in liposome technologies and their applications for systemic
aInex Pharmaceuticals Corporation, 1779 West 75th Street, Vancouver, BC, V6P 6P2, Canada
bDepartment of Biochemistry and Molecular Biology, The University of British Columbia, Vancouver V6T 1Z3, Canada
Received 1 July 1997; accepted 11 July 1997
Abstract
The recent clinical successes experienced by liposomal drug delivery systems stem from the ability to produce
well-defined liposomes that can be composed of a wide variety of lipids, have high drug-trapping efficiencies and have anarrow size distribution, averaging less than 100 nm in diameter. Agents that prolong the circulation lifetime of liposomes,enhance the delivery of liposomal drugs to specific target cells, or enhance the ability of liposomes to deliver drugsintracellularly can be incorporated to further increase the therapeutic activity. The physical and chemical requirements foroptimum liposome drug delivery systems will likely apply to lipid-based gene delivery systems. As a result, the developmentof liposomal delivery systems for systemic gene delivery should follow similar strategies. Keywords: Liposomal gene delivery systems; Cationic lipids; DNA encapsulation; Prolonged circulation lifetimes;Targeting; Fusogenic liposomes; Intracellular delivery; Transfection
Contents
2. Production of liposomes for conventional drug delivery .
3. Production of liposomes for gene delivery.
4. Liposomes with prolonged circulation lifetimes .
5. Targeted delivery to specific cells .
6. Fusogenic liposomes for intracellular delivery . 1. Introduction
al drugs with proven clinical utility [1,2]. Notableexamples are liposome formulations of doxorubicin
Recent advances in liposome technologies for
[3,4], all-trans retinoic acid [5], amphotericin B [6],
conventional drug delivery have resulted in liposom-
daunorubicin [7] and vincristine [8,9]. These ad-vances have led to the production of well-defined,
relatively small liposomal systems that have the
Corresponding author. Tel.: 1 1 604 2649954; fax: 1 1 604
ability to entrap drugs with high efficiencies, reside
1998 Elsevier Science B.V. All rights reserved. P I I S 0 1 6 9 - 4 0 9 X ( 9 7 ) 0 0 1 0 8 - 7
A. Chonn, P.R. Cullis / Advanced Drug Delivery Reviews 30 (1998) 73 –83
in the circulation for extended periods, and accumu-
specific target cells. This review will highlight
late at regional sites of disease, such as inflammation
several of the advances made in liposome tech-
and tumor. The next generation of liposomal drug
nologies and discuss how these advances may be
delivery systems will include drug-loaded liposomes
applied to resolve the challenges facing the develop-
with surface-associated targeting information that
ment of liposomes for the controllable and reproduc-
will increase drug accumulation in specific cells, as
well as fusogenic liposomes that will enable moreefficient intracellular drug delivery.
It is anticipated that these advances in liposome
2. Production of liposomes for conventional
technologies will be directly applicable to the design
drug delivery
of liposomal systems for systemic gene delivery. Inmany ways, the challenges facing the development
The major advances in liposome technology in the
of liposomal gene delivery systems are not unlike
past decade arise from the ability to produce well-
those that have faced liposomal drug delivery sys-
defined liposomes composed of a wide variety of
tems. The therapeutic index of the conventional or
lipids with different physical and chemical prop-
gene-based (plasmid DNA or RNA transcripts) drug
erties, having high drug-trapping efficiencies and
will be enhanced by delivering more biologically
having narrow size distributions, averaging less than
active drug to target cells / tissues and less to non-
100 nm in diameter. These physical and chemical
target cells / tissues, to avoid drug-related toxicities.
properties have been shown to significantly affect the
With gene-based drugs, however, delivery into
stability and pharmacokinetics of liposomes [17]. A
appropriate cells represents only part of the problem;
number of procedures have been established to
a number of intracellular barriers exist that can
produce well-defined liposomes (extensively re-
inhibit the biologic activity of gene-based drugs
viewed in [18,19]). These include extrusion, where
[10,11]. It is not clear what role, if any, liposomes
the liposomes are forced through filters with well-
will play in overcoming these intracellular barriers.
defined pore sizes under moderate pressures, re-
The potential of liposomes to systemically deliver
versed-phase evaporation, sonication and detergent-
DNA was recognized as early as the late 1970s
(extensively reviewed in [12]), however, gene-based
Another significant advance has come from the
drugs have presented interesting challenges for sys-
ability to entrap drugs in liposomes with high
temic delivery systems. First, gene-based drugs are
efficiencies while maintaining the integrity of the
highly susceptible to degradation by the nucleases
liposome structure. Drug loading can be achieved
present in plasma. Although liposomes have the
either passively (i.e. the drug is encapsulated during
potential to encapsulate gene-based drugs and pre-
liposome formation) or actively (i.e. after liposome
vent inactivation by nucleases, procedures to effi-
formation). Hydrophobic drugs can be directly in-
ciently encapsulate plasmid DNA in well defined,
corporated into liposomes during vesicle formation,
small liposomes or lipidic DNA particles have only
and the extent of uptake and retention is governed by
recently been realized. Second, the efficacy of gene-
drug–lipid interactions. Trapping efficiencies of
based drugs is completely dependent on gaining
100% are often achievable, but this is dependent on
entry into the target cell cytosol in an intact form.
the solubility of the drug in the liposome membrane.
Therefore, for liposomes to be effective, they must
Passive encapsulation of water-soluble drugs relies
incorporate agents that promote intracellular deliv-
on the ability of liposomes to trap aqueous buffer
ery. With few exceptions (i.e. skeletal muscle [13,14]
containing a dissolved drug during vesicle formation.
and hepatocytes [15,16]), naked plasmid DNA alone
Trapping efficiencies (generally less than 30%) are
is not taken up very efficiently by most cell types in
limited by the trapped volume contained in the
vivo. Third, for certain gene therapy approaches,
liposomes and drug solubility. Another approach to
such as those involving the delivery of suicide genes,
enhance the passive encapsulation of water-soluble
systemic gene delivery systems must have the po-
drugs is to impart an amphipathic nature to the drugs
tential to selectively deliver gene-based drugs to
by conjugating or complexing the drugs to lipids
A. Chonn, P.R. Cullis / Advanced Drug Delivery Reviews 30 (1998) 73 –83
[20,21]. Alternatively, water-soluble drugs that have
cells in vitro [29] by (1) increasing the association of
ionizable amine functions can be actively entrapped
plasmid DNA with liposomes and (2) increasing the
by employing pH gradients [22], which can result in
binding of cationic liposome–plasmid DNA com-
trapping efficiencies approaching 100%.
plexes to cells. This has prompted many researchersto synthesize different cationic lipids that exhibitimproved gene transfer and cell tolerability prop-
3. Production of liposomes for gene delivery
erties [30–32], as well as to develop novel pro-cedures to efficiently encapsulate plasmid DNA
Based on our experience with liposomal drug
within lipid-based carriers. The addition of plasmid
delivery systems, it is envisioned that the ideal
DNA to preformed cationic liposomes often results
liposomes for systemic gene delivery will encapsu-
in the formation of a heterogeneous mixture of
late plasmid DNA with high efficiencies, will protect
unstable complexes of cationic lipids and plasmid
the DNA from degradation by plasma nucleases, will
DNA [33–35]. This heterogeneity and instability are
have a narrow size distribution, averaging 100 nm or
undoubtedly responsible for the poor reproducibility
less in diameter, in order that the liposomes can
in the transfection activity observed in vivo when
access extravascular regions, and will have the
these complexes are administered intravenously [36–
potential to incorporate a wide range of lipids,
especially lipids that promote fusion with cellular
In the past couple of years, there have been
membranes and / or enhance liposome stability in the
significant advances made in the formulation of
circulation. The feasibility of passively encapsulating
plasmid DNA into relatively small, stable plasmid
DNA in liposomes was demonstrated in the late
DNA-containing lipidic particles or liposomes that
1970s using a number of the methods indicated
protect plasmid DNA from degradation by nucleases.
above. For example, high molecular weight DNA is
For example, Gao and Huang [40] describe a pro-
entrapped in egg phosphatidylcholine liposomes by
cedure where the addition of polylysine or other
hydrating the lipid film in the presence of DNA [23].
polycationic polymers to plasmid DNA, prior to or
In a similar manner, metaphase chromosomes are
during the addition of cationic liposomes, results in
passively entrapped in, or tightly associated with,
particles with membranous structures of less than
egg phosphatidylcholine–cholesterol (7:2, mol / mol)
100 nm in diameter. The plasmid DNA in the
liposomes [24]. Alternatively, DNA can be encapsu-
presence of polylysine alone or polylysine and
lated in cochleate lipid cylinders that are formed
cationic liposomes appears to be resistant to nuclease
from the calcium-induced fusion of phosphatidylser-
attack, remaining supercoiled when incubated with 5
ine liposomes [25]. Reversed-phase evaporation pro-
ml of fetal bovine serum at 378C for 1 h. As
cedures have also been employed to encapsulate
characterized by negative stain electron microscopy,
plasmid DNAs with good but variable encapsulation
however, the plasmid DNA does not appear to be
efficiencies [26,27]. More recently, freeze drying
fully encapsulated by a lipid membrane. Moreover,
methods have yielded DNA-containing multilamellar
the particles formed are heterogeneous in nature,
vesicles with encapsulation efficiencies of 50–60%
[28]. For the most part, however, these procedures
ratios that affect the transfection activity in Chinese
yield relatively large multilamellar vesicles with low
hamster ovary (CHO) cells in vitro. The transfection
DNA encapsulating efficiencies and generally low
active particle can be isolated by sucrose density
gene transfer capabilities. Extrusion of the DNA-
gradient ultracentrifugation and the purified particle
containing multilamellar vesicles to reduce the par-
is reported to be stable for up to three months at 48C,
ticle size have resulted in poor recoveries of DNA-
with no increase in particle size. Similar particles can
be generated by the addition of DOPE, cholesteryl
In the late 1980s, it was shown that cationic lipids,
hemisuccinate and folate–poly(ethylene glycol)–
phosphatidylethanolamine conjugates (6:4:0.01, mol /
ethanolamine (DOPE)-containing liposomes, could
mol / mol) to plasmid DNA–polylysine complexes
enhance the efficiency of gene delivery to cultured
[41]. These particles were shown to be highly active
A. Chonn, P.R. Cullis / Advanced Drug Delivery Reviews 30 (1998) 73 –83
in transfecting KB cells in vitro, being 20–30 times
mid DNA–cationic lipid complex formed by the
more active than 3-b-[N-(N9,N9-dimethylethane)car-
addition of cationic lipids, added in monomer or
bamoyl]cholesterol–DOPE, (6:4, mol / mol) cationic
micellar form, to plasmid DNA [45,46]. This com-
liposome–plasmid DNA complexes. The stability
plex can serve as a well-defined intermediate in the
and pharmacokinetics of these particles upon in-
preparation of plasmid DNA-containing liposomes
travenous administration, or the ability of these
with good properties for systemic gene delivery
particles to transfect cells in vivo, have not been
applications (unpublished results). For example, the
Recently, Hofland et al. [42] described a detergen-
dioleoylphosphatidylcholine or DOPE, to these inter-
t-based procedure to form stable plasmid DNA–
mediates results in the formation of plasmid DNA-
lipidic particles by the addition of various amounts
containing liposomes that have a narrow size dis-
of cationic lipids [2,3-dioleoyloxy-N-(2(sperminecar-
tribution, averaging 70–100 nm in diameter (Fig. 1).
boxamido)-ethyl)-N,N-dimethyl-1-propanaminium tr-
Typically, plasmid DNA encapsulating efficiencies
ifluoroacetate] and DOPE (1.5:1, mol / mol) solubil-
of 70% are obtained using this procedure. A wide
ized in buffered 1% octylglucoside to plasmid DNA,
variety of lipids that alter the biodistribution of the
followed by removal of the detergent by dialysis.
liposomes can be readily incorporated into these
The particles can be stored frozen or as a suspension
liposomes. For example, the incorporation of at least
at 48C for 90 days with no loss in transfection
10 mol% poly(ethylene glycol) conjugated to phos-
activity in NIH 3T3 cells in vitro. The physicalproperties of the active particles have not beendefined. However, the active particles can be pelletedby centrifugation at 3000 g for 15 min, indicatingthat they are relatively large particles. Moreover, invitro transfection efficiency is affected by the pres-ence of serum, with a 70% reduction in transfectionactivity in the presence of as little as 1% fetal bovineserum in the culture medium. Another detergent-based method that has yielded active particles hasrecently been described by Liu et al. [43,44]. In thisprocedure, stable emulsions of cationic lipids andplasmid DNA are produced by the addition of non-ionic surfactants. These particles are not well de-fined, but are relatively large in size. The averagediameter of lipid particles for emulsions containingvarious surfactants range from 170 to 250 nm. Uponmixing with plasmid DNA, the particle size increasesfive- to fourteen-fold in diameter, depending on thetype of non-ionic surfactant used for preparing theemulsions. The use of detergents containing branch-ed polyoxyethylene chains as the hydrophilic headgroup are more effective in preventing the formationof large DNA–emulsion complexes. The stabilityand biodistribution of these particles upon intraven-ous administration, or the ability of these particles totransfect cells in vivo have not yet been described inthe literature.
Fig. 1. Cryo-electron micrographs of plasmid DNA encapsulated
An alternative approach that has recently been
in liposomes. Panel (A) represents vesicles formed in the presence
developed takes advantage of the hydrophobic plas-
of and (B) in the absence of plasmid DNA. A. Chonn, P.R. Cullis / Advanced Drug Delivery Reviews 30 (1998) 73 –83
phatidylethanolamine results in plasmid DNA-con-
the target cell of several gene therapies for blood
taining liposomes that have a circulation half-life
approaching 10–12 h in mice. Moreover, these
The biodistribution of intravenously administered
DNA-containing liposomes appear to be stable in the
cationic liposome–plasmid DNA complexes is not
circulation of mice, with the majority of the plasmid
appropriate for such systemic applications. For in-
DNA extracted from the circulating liposomes at 24
stance, it has recently been demonstrated that cat-
ionic liposome–plasmid DNA complexes, exhibitingstrong positive zeta potentials, are cleared rapidlyfrom the circulation [57,58]. These intravenously
4. Liposomes with prolonged circulation
administered cationic liposome–plasmid DNA com-
lifetimes
plexes [N-(2,3-bis(oleyloxy)propyl-N,N,N-trimethyl-ammonium
The use of liposomes for systemic drug delivery
monium bromide and DOPE-containing liposomes]
requires that the liposomes have the ability to avoid
are rapidly eliminated from the plasma, with 50–
immediate uptake by phagocytic cells of the re-
60% of the dose taken up by the liver within 5 min,
ticuloendothelial system (RES) and remain in circu-
and 20–30% of the dose taken up by the lung within
lation for extended periods of time in order to
1 min, falling to 10% after 5 min [57]. The cationic
enhance the opportunity for the liposomal drugs to
liposome–plasmid DNA complexes are predominant-
reach non-RES target tissues. A significant advance
ly taken up by the Kupffer cells in the liver.
in the development of liposomal drugs has come
Moreover, a recent study has shown that cationic
with the use of specialized lipids, such as mono-
lipid–DNA complexes, harboring excess positive
surface charge, are potent activators of the comple-
)-modified phosphatidylethanolamine, that engender
ment system, potentially a barrier to the efficient
long circulation lifetimes when incorporated into
delivery of genes when using high lipid doses [59].
liposomes [47–50]. It has been proposed that these
Although there have been a few reports demon-
PEG–lipid conjugates provide a ‘steric stabilization’
strating the feasibility of using these complexes to
of the surface by virtue of the hydrophilic brush coat
deliver genes to a number of different tissues (such
provided by the PEG polymer [51]. This coat has
as the liver, lung, spleen, heart, skeletal muscle,
been shown to inhibit serum protein binding to the
kidney, uterus, bone marrow cells, peripheral blood
liposomal surface [52,53], which would otherwise
and ovary) after intravenous administration [36–
promote uptake by the RES, complement activation
39,57,58], the observed levels of gene delivery are
and destabilization of the liposomal membranes. It
low and often are not reproducible. This may be a
has been demonstrated that increased circulation
consequence of the rapid elimination of the majority
lifetimes enhance the opportunity for liposomes,
of the injected dose of cationic liposome–plasmid
administered systemically, to leave the vascular
compartment and enter certain extravascular regions
Our recent findings show that DOPE-containing
cationic liposomes can be stabilized in the circulation
The ability to generate sustained circulating
of mice by reducing the cationic lipid content of the
liposomal gene delivery systems using the PEG–
liposomes and incorporating at least 2 mol% PEG–
lipid technology should prove useful for systemic
phosphatidylethanolamine derivatives [60]. In vitro,
gene delivery applications. For instance, the ability
the addition of serum to cationic liposomes com-
of long circulating liposomes to accumulate within
tumors will be advantageous for cancer gene therapy
DOPE (85:15, mol / mol) induces a rapid aggregation
applications involving tumor suppressor genes or
of the cationic liposomes, forming large fused aggre-
suicide genes. Furthermore, the avoidance of RES
gates ( . 1 mm in diameter) [60]. Amphipathic
uptake, especially by Kupffer cells, the resident
PEG–lipid conjugates can stabilize DOPE-containing
macrophages of the liver, would enhance the oppor-
liposomes by inhibiting the fusogenic activity of
tunity for liposomes to deliver genes to hepatocytes,
A. Chonn, P.R. Cullis / Advanced Drug Delivery Reviews 30 (1998) 73 –83
However, the fusogenic activity is essential for
DNA-containing liposomes has recently been dem-
efficient gene delivery [30,31,41,63,64] and, thus, an
onstrated in vitro by a number of investigators. For
essential property of the amphipathic PEG–lipid
example, Lee and Huang [41] have shown that
conjugates is that they have the ability to dissociate
folate, conjugated to the distal end of PEG–phos-
from the carrier at some later time, restoring the
phatidylethanolamine, enhances the plasmid DNA
fusogenic activity of the liposomes and allowing the
uptake and transfection efficiency of KB cells in
liposomes to fuse with target cells. The feasibility of
vitro by employing plasmid DNA-containing pH-
this approach has recently been demonstrated [61].
The rate at which fusogenic activity is recovered is
mol / mol) liposomes, particularly when the lipo-
shown to be controlled to a large extent by the same
somes carry an overall negative surface charge. This
parameters that regulate spontaneous transfer of
study clearly demonstrates that components which
enhance the binding of liposomes to cells, mediatedeither by the use of targeting ligands or by a strongpositive surface charge, are essential for efficient
5. Targeted delivery to specific cells
liposomal gene delivery systems. The addition oftransferrin to cationic liposome–plasmid DNA com-
In general, liposomes are effective delivery sys-
plexes increases the amount of DNA taken up by
tems because they alter the pharmacokinetics of the
human hepatoma HepG2 cells in vitro twofold,
free drug, leading to enhanced drug bioavailability to
accompanied by a significant increase in the number
specific target cells that reside in the circulation or,
of b-galactosidase-positive cells (98–100% in the
more importantly, to extravascular disease regions.
presence of transferrin compared to 3–4% in the
The ability to selectively deliver drugs to specific
absence of transferrin) [83]. Transferrin presumably
cells, such as tumor cells, within these regions will
acts to further facilitate the uptake of cationic
further enhance the therapeutic index of liposomal
liposome–DNA complexes via a receptor-mediated
drugs. Targeted delivery and improved therapeutic
process. Similarly, asialofetuin [79,80] and galac-
activity of liposomal drugs in vivo has been achieved
tose-containing lipids [84] have been shown to
by coupling site-directive targeting ligands, such as
increase the transfection efficiency of HepG2 cells in
monoclonal antibodies [65–68], to the surface of
vitro. Kikuchi et al. [85] have shown that the
liposomes by either covalent or non-covalent meth-
addition of epidermal growth factor to cationic
ods [68,69]. A significant advance in this area has
liposomes enhances the in vitro luciferase gene
been the advent of novel PEG–phosphatidylethanol-
expression in epidermal growth factor receptor-over-
amine lipids that allow targeting ligands to be
expressing HEC-A cells and not in epidermal growth
conjugated at the distal ends of the PEG spacer
factor receptor-deficient HRA cells. The coupling of
[70–73]. These conjugates increase target cell bind-
antibodies to pH-sensitive liposomes [86] or to
ing in vitro, as well as prolong circulation times.
cationic liposomes [87] has been shown to also
Furthermore, in addition to antibodies, glycolipids
enhance transfection activity in vitro compared to
[74–77], proteins [78–80] and vitamins [41,71] have
that found in non-targeted DNA-containing lipo-
been used to selectively target specific cells via cell
For liposomal gene delivery systems, targeting
ligands need to function not only to increase the
6. Fusogenic liposomes for intracellular delivery
binding of the liposomes to specific target cells, suchas hepatocytes, but also to promote the cellular
Fusogenic liposomes can potentially facilitate the
uptake of the liposomes via an endocytic pathway.
intracellular delivery of encapsulated drugs by fusing
Endocytosis is believed to play a major role in
with the target cell. A variety of approaches can be
plasmid DNA delivery to cultured cells in vitro
envisioned for constructing fusogenic liposomes.
[10,63,81,82]. The feasibility of using targeting
Examples include the inclusion of lipids that are able
ligands to increase the cellular uptake of plasmid
to form non-bilayer phases, such as DOPE, which
A. Chonn, P.R. Cullis / Advanced Drug Delivery Reviews 30 (1998) 73 –83
can promote destabilization of the bilayer, inducing
propane) can function in the absence of helper lipids
fusion events [88,89]. Furthermore, alterations in the
[30,31] or in the presence of cholesterol (such as
lipid composition can render liposomes pH sensitive,
dioctadecylammonium bromide) [38] suggests that
leading to enhanced fusogenic tendencies in low pH
these cationic lipids may, by themselves, possess
compartments such as endosomes [41,86,90]. Non-
properties that promote endosomal release of plas-
phospholipid fusogenic liposomes composed primari-
mids via a mechanism other than a membrane fusion
ly of dioxyethylene acyl ethers and cholesterol have
event. As previously mentioned, plasmid DNA-con-
been shown to fuse with plasma membranes of
taining pH-sensitive liposomes are efficient gene
erythrocytes and fibroblasts [91]. Alternatively, effi-
delivery systems in vitro, provided that they have
cient fusogenic liposomes can be achieved by incor-
targeting ligands coupled to their surface [41,86].
porating fusogenic proteins into the liposome mem-brane [92–94] or entrapped within liposomes [95]. The feasibility of this approach has been demon-
7. Conclusions
strated for the delivery of the diphtheria toxin Asubunit using liposomes produced from influenza
The development of controllable and reproducible
virus envelopes [94]. Fusogenic peptides can be
liposomal systems for systemic gene delivery neces-
conjugated to the liposomes [96–98] and may also
sitates the establishment of methods to efficiently
promote intracellular delivery. The encapsulation of
encapsulate gene-based drugs in well-defined, rela-
a 30-amino acid fusogenic peptide has recently been
tively small liposomes. Traditional methods for
shown to promote relatively efficient endosomal
encapsulating drugs in liposomes have proven to be
release of propidium iodide, with 20–25% of the
ineffective for gene-based drugs. However, recently
encapsulated propidium iodide gaining access to KB
developed detergent-based procedures to produce
cell chromosomal DNA after 48 h [99].
stable plasmid DNA lipidic particles or plasmid
The effectiveness of liposomal gene-based drugs is
DNA-containing liposomes appear promising. In
dependent on their ability to access the cytosol of
vitro studies have shown that these systems are
target cells. For optimum efficiency, therefore, lipid-
active in delivering plasmid DNA to a number of
based gene delivery systems should exhibit fusogenic
cultured established cell lines. The in vivo studies
activity. A number of studies illustrate that the above
are certainly forthcoming. Although these are early
approaches to enhance the fusogenic activity of
stages for liposomal gene delivery systems, several
liposomes can be applied to enhance the efficiency of
of the advances made in liposomal drug delivery
lipid-based gene delivery systems. For example, the
technologies can be directly applied to these systems.
addition of replication-deficient adenovirus, which
Noteworthy is the use of exchangeable PEG–lipid
enhance endosomal escape, to cationic liposome–
conjugates to stabilize the plasmid DNA-containing
plasmid DNA complexes results in an approximately
lipid-based carriers in the circulation. This should
fivefold increase in chloramphenicol acyl transferase
expedite the development of systemic liposomal gene
activity detected in FAO hepatoma and 3T3-F442A
delivery systems that exhibit targeted and enhanced
adipocyte cells in vitro [100], and up to a 1000-fold
increase in luciferase expression in human smoothmuscle cells in vitro [101]. Similarly, the incorpora-tion of the fusogenic protein from Sendai virus, by
References
fusing Sendai virus with preformed DNA-containingliposomes, results in a liposome with improved gene
[1] A. Chonn, P.R. Cullis, Recent advances in liposomal drug-
delivery properties [102–104]. For the majority of
delivery systems, Curr. Opin. Biotechnol. 6 (1995) 698–708.
[2] G. Gregoriadis, Engineering liposomes for drug delivery:
cationic lipids, DOPE is required for efficient gene
progress and problems, TIBTECH 13 (1995) 527–537.
delivery to cells in vitro [30,31,63,64]. However, the
[3] G.K. Schwartz, E.S. Casper, A phase II trial of doxorubicin
observation that certain cationic lipids (such as
HCl liposome injection in patients with advanced pancreatic
adenocarcinoma, Invest. New Drugs 13 (1995) 77–82.
[4] J. Treat, A. Greenspan, D. Forst, J.A. Sanchez, V.J. Ferrans,
A. Chonn, P.R. Cullis / Advanced Drug Delivery Reviews 30 (1998) 73 –83
L.A. Potkul, P.V. Woolley, A. Rahman, Antitumor activity of
[19] R.M. Watwe, J.R. Bellare, Manufacture of liposomes: a
liposome-encapsulated doxorubicin in advanced breast can-
review, Curr. Sci. 68 (1995) 715–724.
cer: phase II study, J. Natl. Cancer Inst. 82 (1990) 1706–
[20] M. van Borssum-Waalkes, M. van Galen, H. Morselt, B.
Sternberg, G.L. Scherphof, In vitro stability and cytostatic
[5] E. Estey, P.F. Thall, K. Mehta, M. Rosenblum, T. Brewer Jr.,
activity of liposomal formulations of 5-fluoro-29-deox-
V. Simmons, F. Cabanillas, R. Kurzrock, G. Lopez-Berestein,
yuridine and its diacylated derivatives, Biochim. Biophys.
Alterations in tretinoin pharmacokinetics following adminis-
tration of liposomal all-trans-retinoic acid, Blood 87 (1996)
[21] O. Zelphati, E. Wagner, L. Leserman, Synthesis and anti-HIV
activity of thiocholesteryl-coupled phosphodiester antisense
[6] K.M. Wasan, G. Lopez-Berestein, The past, present, and
oligonucleotides incorporated into immunoliposomes, An-
future uses of liposomes in treating infectious diseases,
Immunopharmacol. Immunotoxicol. 17 (1995) 1–15.
[22] L.D. Mayer, T.M. Madden, M.B. Bally, P.R. Cullis, pH
[7] P.S. Gill, J. Wernz, D.T. Scadden, P. Cohen, G.M. Mukwaya,
gradient-mediated drug entrapment in liposomes, in: G.
J.H. von Roenn, M. Jacobs, S. Kempin, I. Silverberg, G.
Gregoriadis, (Ed.), Liposome Technology, Vol. 2, 2nd edi-
Gonzales et al., Randomized phase III trial of liposomal
tion, CRC Press, Boca Raton, FL, 1993, pp. 27–44.
daunorubicin versus doxorubicin, bleomycin, and vincristine
[23] R.M. Hoffman, L.B. Margolis, L.D. Bergelson, Binding and
in AIDS-related Kaposi’s sarcoma, J. Clin. Oncol. 14 (1996)
entrapment of high molecular weight DNA by lecithin
liposomes, FEBS Lett. 93 (1978) 365–368.
[8] L.D. Mayer, D. Masin, R. Nayar, N.L. Boman, M.B. Bally,
[24] A.B. Mukherjee, S. Orloff, J.D. Butler, T. Triche, P. Lalley,
Pharmacology of liposomal vincristine in mice bearing
J.D. Schulman, Entrapment of metaphase chromosomes into
L1210 ascitic and B16 / BL6 solid tumours, Br. J. Cancer 71
phospholipid vesicles (lipochromosomes): carrier potential in
gene transfer, Proc. Natl. Acad. Sci. U.S.A. 75 (1978)
[9] M.S. Webb, T.O. Harasym, D. Masin, M.B. Bally, L.D.
Mayer, Sphingomyelin–cholesterol liposomes significantly
[25] R.J. Mannino, E.S. Allebach, W.A. Strohl, Encapsulation of
enhance the pharmacokinetic and therapeutic properties of
high molecular weight DNA in large unilamellar phos-
vincristine in murine and human tumour models, Br. J.
pholipid vesicles, FEBS Lett. 101 (1979) 229–232.
[26] M. Schaefer-Ridder, Y. Wang, P.H. Hofschneider, Liposomes
[10] J. Zabner, A.J. Fasbender, T. Moninger, K.A. Peollinger,
as gene carriers: efficient transformation of mouse L cells by
M.J. Welsh, Cellular and molecular barriers to gene transfer
thymidine kinase gene, Science 215 (1982) 166–168.
by a cationic lipid, J. Biol. Chem. 270 (1995) 18997–19007.
[27] C. Nicolau, A. Le Pape, P. Soriano, F. Fargette, M.F. Juhel,
[11] M.E. Dowty, P. Williams, G. Zhang, J.E. Hagstrom, J.A.
In vivo expression of rat insulin after intravenous administra-
Wolff, Plasmid DNA entry into postmitotic nuclei of primary
tion of the liposome-entrapped gene for rat insulin I, Proc.
rat myotubes, Proc. Natl. Acad. Sci. U.S.A. 92 (1995)
Natl. Acad. Sci. U.S.A. 80 (1983) 1068–1072.
[28] M. Baru, J.H. Axelrod, I. Nur, Liposome-encapsulated DNA-
[12] P. Hug, R.G. Sleight, Liposomes for the transformation of
mediated gene transfer and synthesis of human factor IX in
eukaryotic cells, Biochim. Biophys. Acta 1097 (1991) 1–17.
[13] J.A. Wolff, R.W. Malone, P. Williams, W. Chong, G. Ascadi,
A. Jani, P.L. Felgner, Direct gene transfer into mouse muscle
[29] P.L. Felgner, T.R. Gadek, M. Holm, R. Roman, H.W. Chan,
in vivo, Science 247 (1990) 1465–1468.
M. Wenz, J.P. Northrop, G.M. Ringold, M. Danielsen,
[14] J. Wolff, M. Dowty, S. Jiao, G. Repetto, R. Berg, J. Ludtke,
Lipofection: a highly efficient, lipid-mediated DNA transfec-
P. Williams, Expression of naked plasmids by cultured
tion procedure, Proc. Natl. Acad. Sci. U.S.A. 84 (1987)
myotubes and entry of plasmids into T tubules and caveolae
of mammalian skeletal muscle, J. Cell Sci. 103 (1992)
[30] J.H. Felgner, R. Kumar, C.N. Sridhar, C.J. Wheeler, Y.J.
Tsai, R. Border, P. Ramsey, M. Martin, P.L. Felgner,
[15] M. Yoshida, R.I. Mahato, K. Kawabata, Y. Takakura, M.
Enhanced gene delivery and mechanism studies with a novel
Hashida, Disposition characteristics of plasmid DNA in the
series of cationic lipid formulations, J. Biol. Chem. 269
single pass rat liver perfusion system, Pharm. Res. 13 (1996)
[31] X. Gao, L. Huang, Cationic liposome-mediated gene trans-
[16] M.A. Hickman, R.W. Malone, K. Lehmann-Bruinsma, T.R.
Sih, D. Knoell, F.C. Szoka Jr., R. Walzem, D.M. Carlson,
[32] R.P. Balasubramaniam, M.J. Bennett, A.M. Aberle, J.G.
J.S. Powell, Gene expression following direct injection of
Malone, M.H. Nantz, R.W. Malone, Structural and functional
DNA into liver, Hum. Gene Ther. 5 (1994) 1477–1483.
analysis of cationic transfection lipids: the hydrophobic
[17] J. Senior, Fate and behavior of liposomes in vivo: a review
domain, Gene Ther. 3 (1996) 163–172.
of controlling factors, Crit. Rev. Ther. Drug Carrier Syst. 3
[33] H. Gershon, R. Ghirlando, S.B. Guttman, A. Minsky, Mode
of formation and structural features of DNA–cationic lipo-
[18] G. Gregoriadis, Liposome preparation and related tech-
some complexes used for transfection, Biochemistry 32
niques, in: G. Gregoriadis, (Ed.), Liposome Technology, Vol.
1, 2nd edition, CRC Press, Boca Raton, FL, 1993, pp. 1–63.
[34] J. Gustafsson, G. Arvidson, G. Karlsson, M. Almgren,
A. Chonn, P.R. Cullis / Advanced Drug Delivery Reviews 30 (1998) 73 –83
Complexes between cationic liposomes and DNA visualized
therapeutic efficacy, Proc. Natl. Acad. Sci. U.S.A. 88 (1991)
by cryo-TEM, Biochim. Biophys. Acta 1235 (1995) 305–
[51] D. Needham, T.J. McIntosh, D.D. Lasic, Repulsive interac-
[35] B. Sternberg, F.L. Sorgi, L. Huang, New structures in
tions and mechanical stability of polymer-grafted lipid
complex formation between DNA and cationic liposomes
membranes, Biochim. Biophys. Acta 1108 (1992) 40–48.
visualized by freeze-fracture electron microscopy, FEBS
[52] S. Semple, A. Chonn, Protein–liposome interactions in
relation to clearance, J. Liposome Res. 6 (1996) 33–60.
[36] D. Lew, S.E. Parker, T. Latimer, A.M. Abai, A. Kuwahara-
[53] J. Senior, C. Delgado, D. Fisher, C. Tilcock, G. Gregoriadis,
Rundell, S.G. Doh, Z. Yang, D. Laface, S.H. Gromkowski,
Influence of surface hydrophilicity of liposomes on their
G.J. Nabel, M. Manthorpe, J. Norman, Cancer gene therapy
interaction with plasma protein and clearance from the
using plasmid DNA: pharmacokinetic study of DNA follow-
circulation: studies with poly(ethylene glycol)-coated vesi-
ing injection in mice, Hum. Gene Ther. 6 (1995) 553–564.
cles, Biochim. Biophys. Acta 1062 (1991) 77–82.
[54] A. Gabizon, D. Papahadjopoulos, Liposome formulations
Rabinovich, R.C. Gallo, L.C. Mahan, Systemic gene therapy:
with prolonged circulation time in blood and enhanced
biodistribution and long-term expression of a transgene in
uptake by tumors, Proc. Natl. Acad. Sci. U.S.A. 85 (1988)
mice, Proc. Natl. Acad. Sci. U.S.A. 92 (1995) 9742–9746.
[38] Y. Liu, D. Liggitt, W. Zhong, G. Tu, K. Gaensler, R. Debs,
[55] I.A.J.M. Bakker-Woundenberg, A.F. Lokerse, M.T. Ten-
Cationic liposome-mediated intravenous gene delivery, J.
Kate, G. Storm, Enhanced localization of liposomes with
Biol. Chem. 270 (1995) 24864–24870.
prolonged blood circulation time in infected lung tissue,
[39] N. Zhu, D. Liggitt, Y. Liu, R. Debs, Systemic gene expres-
Biochim. Biophys. Acta 1138 (1992) 318–326.
sion after intravenous DNA delivery into adult mice, Science
[56] N.Z. Wu, D. Da, T.L. Rudolf, D. Needham, A.R. Whorton,
M.W. Dewhirst, Increased microvascular permeability contri-
[40] X. Gao, L. Huang, Potentiation of cationic liposome-me-
butes to preferential accumulation of stealth liposomes in
diated gene delivery by polycations, Biochemistry 35 (1996)
tumor tissue, Cancer Res. 53 (1993) 3765–3770.
[57] R.I. Mahato, K. Kawabata, T. Nomura, Y. Takakura, M.
[41] R.J. Lee, L. Huang, Folate-targeted, anionic liposome-en-
Hashida, Physicochemical and pharmacokinetic characteris-
trapped polylysine-condensed DNA for tumor cell-specific
tics of plasmid DNA / cationic liposome complexes, J.
gene transfer, J. Biol. Chem. 271 (1996) 8481–8487.
[42] H.E. Hofland, L. Shephard, S.M. Sullivan, Formation of
[58] G. Osaka, K. Carey, A. Cuthbertson, P. Godowski, T.
stable cationic lipid / DNA complexes for gene transfer, Proc.
Patapoff, A. Ryan, T. Gadek, J. Mordenti, Pharmacokinetics,
Natl. Acad. Sci. U.S.A. 93 (1996) 7305–7309.
tissue distribution, and expression efficiency of plasmid [P-
[43] F. Liu, J. Yang, L. Huang, D. Liu, New cationic lipid
33]DNA following intravenous administration of DNA / cat-
formulations for gene transfer, Pharm. Res. (1997), in press.
ionic lipid complexes in mice: use of a novel radionuclide
[44] F. Liu, J. Yang, L. Huang, D. Liu, Effect of non-ionic
approach, J. Pharm. Sci. 85 (1996) 612–618.
surfactants on the formation of DNA / emulsion complex and
[59] C. Plank, K. Mechtler, F.C. Szoka Jr., E. Wagner, Activation
the emulsion-mediated gene transfer, Pharm. Res. (1997), in
of the complement system by synthetic DNA complexes: a
potential barrier for intravenous gene delivery, Hum. Gene
[45] D.L. Reimer, Y. Zhang, S. Kong, J.J. Wheeler, R.W. Graham,
M.B. Bally, Formation of novel hydrophobic complexes
[60] A. Mori, A. Chonn, L.S. Choi, A. Israels, M.A. Monck, P.R.
between cationic lipids and plasmid DNA, Biochemistry 34
Cullis, In vivo stabilization and regulated fusion of lipo-
somes containing a cationic lipid using amphipathic poly-
[46] F.M.P. Wong, D.L. Reimer, M.B. Bally, Cationic lipid
ethyleneglycol derivatives, submitted.
binding to DNA: characterization of complex formation,
[61] J.W. Holland, C. Hui, P.R. Cullis, T.D. Madden, Poly-
(ethylene glycol)–lipid conjugates regulate the calcium-in-
[47] T.M. Allen, A. Chonn, Large unilamellar liposomes with low
duced fusion of liposomes composed of phosphatidylethanol-
uptake into the reticuloendothelial system, FEBS Lett. 223
amine and phosphatidylserine, Biochemistry 35 (1996)
[48] A.L. Klibanov, K. Maruyama, V.P. Torchilin, L. Huang,
[62] J.W. Holland, P.R. Cullis, T.D. Madden, Poly(ethylene gly-
Amphipathic polyethyleneglycols effectively prolong the
col)–lipid conjugates promote bilayer formation in mixtures
circulation time of liposomes, FEBS Lett. 268 (1990) 235–
of non-bilayer-forming lipids, Biochemistry 35 (1996) 2610–
[49] G. Blume, G. Cevc, Liposomes for the sustained drug release
[63] X. Zhou, L. Huang, DNA transfection mediated by cationic
in vivo, Biochim. Biophys. Acta 1029 (1990) 91–97.
liposomes containing lipopolylysine: characterization and
[50] D. Papahadjopoulos, T.M. Allen, A. Gabizon, E. Mayhew,
mechanism of action, Biochim. Biophys. Acta 1189 (1994)
K. Matthay, S.K. Huang, K.D. Lee, M.C. Woodle, D.D.
Lasic, C. Redemann, F.J. Martin, Sterically stabilized lipo-
[64] H. Farhood, N. Serbina, L. Huang, The role of dioleoyl-
somes: improvements in pharmacokinetics and antitumor
phosphatidylethanolamine in cationic liposome mediated
A. Chonn, P.R. Cullis / Advanced Drug Delivery Reviews 30 (1998) 73 –83
gene transfer, Biochim. Biophys. Acta 1235 (1995) 289–
Technology, Vol. 3, 2nd edition, CRC Press, Boca Raton, FL,
[65] I. Ahmad, M. Longenecker, J. Samuel, T.M. Allen,
[78] J.C. Stavridis, G. Deliconstantinos, M.C. Psallidopoulos,
Antibody-targeted delivery of doxorubicin entrapped in
N.A. Armenakas, D.J. Hadjiminas, J. Hadjiminas, Construc-
sterically stabilized liposomes can eradicate lung cancer in
tion of transferrin-coated liposomes for in vivo transport of
mice, Cancer Res. 53 (1993) 1484–1488.
exogenous DNA to bone marrow erythroblasts in rabbits,
[66] A. Mori, S.J. Kennel, M. Van Borssum-Waalkes, G.L.
Exp. Cell Res. 164 (1986) 568–572.
Scherphof, L. Huang, Characterization of organ-specific
[79] T. Hara, Y. Aramaki, S. Takada, K. Koike, S. Tsuchiya,
immunoliposomes for delivery of 39,59-O-dipalmitoyl-5-
Receptor-mediated transfer of pSV2CAT DNA to a human
fluoro-29-deoxyuridine in a mouse lung-metastasis model,
hepatoblastoma cell line HepG2 using asialofetuin-labeled
Cancer Chemother. Pharmacol. 35 (1995) 447–456.
cationic liposomes, Gene 159 (1995) 167–174.
[67] J.W. Park, K. Hong, P. Carter, H. Asgari, L.Y. Guo, G.A.
[80] T. Hara, H. Kuwasawa, Y. Aramaki, S. Takada, K. Koike, K.
Keller, C. Wirth, R. Shalaby, C. Kotts, W.I. Wood et al.,
Ishidate, H. Kato, S. Tsuchiya, Effects of fusogenic and
DNA-binding amphiphilic compounds on the receptor-me-
therapy, Proc. Natl. Acad. Sci. U.S.A. 92 (1995) 1327–1331.
diated gene transfer into hepatic cells by asialofetuin-labeled
[68] T.M. Allen, Long-circulating (sterically stabilized) liposomes
liposomes, Biochim. Biophys. Acta 1278 (1996) 51–58.
for targeted drug delivery, Trends Pharmacol. Sci. 15 (1994)
[81] I. Wrobel, D. Collins, Fusion of cationic liposomes with
mammalian cells occurs after endocytosis, Biochim. Bio-
[69] H.C. Loughrey, L.S. Choi, K.F. Wong, P.R. Cullis, M.B.
Bally, Preparation of streptavidin–liposomes for use in
[82] F. Labatmoleur, A.M. Steffan, C. Brisson, H. Perron, O.
ligand specific targeting applications, in G. Gregoriadis,
Feugeas, P. Furstenberger, F. Oberling, E. Brambilla, J.P.
(Ed.), Liposome Technology, Vol. 3, 2nd edition, CRC Press,
Behr, An electron microscopy study into the mechanism of
Boca Raton, FL, 1993, pp. 163–178.
gene transfer with lipopolyamines, Gene Ther. 3 (1996)
[70] G. Blume, G. Cevc, M.D.J.A. Crommelin, I.A.J.M. Bakker-
Woudenberg, C. Kluft, G. Storm, Specific targeting with
[83] P. Cheng, Receptor ligand-facilitated gene transfer: enhance-
poly(ethylene glycol)-modified liposomes: coupling of hom-
ment of liposome-mediated gene transfer and expression by
ing devices to the ends of the polymeric chains combines
transferrin, Hum. Gene Ther. 7 (1996) 275–282.
effective target binding with long circulation times, Biochim.
[84] J. Remy, A. Kichler, V. Mordvinov, F. Schuber, J. Behr,
Biophys. Acta 1149 (1993) 180–184.
[71] R.J. Lee, P.S. Low, Delivery of liposomes into cultured KB
lipopolyamine-condensed DNA particles presenting galac-
cells via folate receptor-mediated endocytosis, J. Biol. Chem.
tose ligands: a stage toward artificial viruses, Proc. Natl.
Acad. Sci. U.S.A. 92 (1995) 1744–1748.
[72] K. Maruyama, T. Takizawa, T. Yuda, S.J. Kennel, L. Huang,
[85] A. Kikuchi, S. Sugaya, H. Ueda, K. Tanaka, Y. Aramaki, T.
M. Iwatsuru, Targetability of novel immunoliposomes modi-
Hara, H. Arima, S. Tsuchiya, T. Fuwa, Efficient gene
fied with amphipathic poly(ethylene glycols) conjugated at
transfer to EGF receptor overexpressing cancer cells by
their distal terminals to monoclonal antibodies, Biochim.
means of EGF-labeled cationic liposomes, Biochim. Bio-
phys. Res. Commun. 227 (1996) 666–671.
[73] T.M. Allen, E. Brandeis, C.B. Hansen, G.Y. Kao, S.
[86] C.Y. Wang, L. Huang, Highly efficient DNA delivery me-
Zalipsky, A new strategy for attachment of antibodies to
diated by pH-sensitive immunoliposomes, Biochemistry 28
sterically stabilized liposomes resulting in efficient targeting
to cancer cells, Biochim. Biophys. Acta 1237 (1995) 99–
[87] G.Y. Kao, L.J. Chang, T.M. Allen, Use of targeted cationic
liposomes in enhanced DNA delivery to cancer cells, Cancer
[74] A. Sasaki, N. Murahashi, H. Yamada, A. Morikawa, Syn-
thesis of novel galactosyl ligands for liposomes and the
[88] H. Ellens, J. Bentz, F.C. Szoka Jr., Destabilization of
influence of the spacer on accumulation in the rat liver, Biol.
phosphatidylethanolamine liposomes at the hexagonal phase
transition temperature, Biochemistry 25 (1986) 285–294.
[75] M. Blom, L. Andersson, A. Carlsson, B. Herslof, L. Zhou, A.
[89] H. Ellens, J. Bentz, F.C. Szoka Jr., Fusion of phosphatidyl-
Nilsson, Pharmacokinetics, tissue distribution and metabo-
ethanolamine-containing liposomes and mechanism of the
lism of intravenously administered digalactosyldiacylglycerol
phase transition, Biochemistry 25 (1986) 4141–
and monogalactosyldiacylglycerol in the rat, J. Liposome
[90] A.M. Tari, F. Zhou, L. Huang, Two types of pH-sensitive
[76] H. Yamauchi, H. Kikuchi, M. Sawada, M. Tomikawa, S.
immunoliposomes, in G. Gregoriadis, (Ed.), Liposome Tech-
Hirota, Selective uptake of liposomes containing lactose
nology, Vol. 3, 2nd edition, CRC Press, Boca Raton, FL,
mono-fatty acid derivatives by hepatic parenchymal cells, J.
Microencapsulation 11 (1994) 287–296.
[91] M.E. Baraka, E.I. Pecheur, D.F.H. Wallach, J.R. Philippot,
[77] G. Barratt, F. Schuber, Targeting of liposomes with man-
Non-phospholipid fusogenic liposomes, Biochim. Biophys.
nose-terminated ligands, in: G. Gregoriadis, (Ed.), Liposome
A. Chonn, P.R. Cullis / Advanced Drug Delivery Reviews 30 (1998) 73 –83
[92] B. Compagnon, P. Milhaud, A. Bienvenue, J.R. Philippot,
bound to liposomes, Biochim. Biophys. Acta 1195 (1994)
Targeting of poly(rI)–poly(rC) by fusogenic (F proteins)
immunoliposomes, Exp. Cell Res. 200 (1992) 333–338.
[99] K. Vogel, S. Wang, R.J. Lee, P.S. Low, J. Chmielewski,
[93] C.M. de Fiebre, S.O. Bryant, D. Notabartolo, P. Wu, E.M.
Peptide-mediated release of folate-targeted liposome contents
Meyer, Fusogenic properties of Sendai virosome envelopes
from endosomal compartments, J. Am. Chem. Soc. (1996).
in rat brain preparations, Neurochem. Res. 18 (1993) 1089–
[100] C. Meunier-Durmort, N. Ferry, B. Hainque, J. Delattre, C.
Forest, Efficient transfer of regulated genes in adipocytes
[94] R. Bron, A. Ortiz, J. Wilschut, Cellular cytoplasmic delivery
and hepatoma cells by the combination of liposomes and
of a polypeptide toxin by reconstituted Influenza virus
replication-deficient adenovirus, Eur. J. Biochem. 237
envelopes (virosomes), Biochemistry 33 (1994) 9110–9117.
[95] S.E. Glushakova, V.G. Omelyanenko, I.S. Lukashevitch, J.
[101] J. Kreuzer, S. Denger, F. Reifers, C. Beisel, K. Haack, J.
Bogdanov, A.B. Moshnikova, A.T. Kozytch, V.P. Torchilin,
Gebert, W. Kubler, Adenovirus-assisted lipofection: effi-
The fusion of artificial lipid membranes induced by the
cient in vitro gene transfer of luciferase and cytosine
synthetic arenavirus ‘fusion peptide’, Biochim. Biophys.
deaminase to human smooth muscle cells, Atherosclerosis
[96] K. Lee, Y. Oh, D.A. Portnoy, J.A. Swanson, Delivery of
[102] Y. Kaneda, K. Iwai, T. Uchida, Increased expression of
macromolecules into cytosol using liposomes containing
DNA cointroduced with nuclear protein in adult rat liver,
hemolysin from Listeria monocytogenes, J. Biol. Chem. 271
[103] I. Yanagihara, K. Inui, G. Dickson, G. Turner, T. Piper, Y.
[97] T. Kato, S. Lee, S. Ono, Y. Agawa, H. Aoyagi, M. Ohno, N.
Kaneda, S. Okada, Expression of full-length human
Nishino, Conformational studies of amphipathic a-helical
dystrophin cDNA in mdx mouse muscle by HVJ–liposome
peptides containing an amino acid with a long alkyl chain
injection, Gene Ther. 3 (1996) 549–553.
and their anchoring to lipid bilayer liposomes, Biochim.
[104] V.J. Dzau, M.J. Mann, R. Morishita, Y. Kaneda, Fusigenic
Biophys. Acta 1062 (1991) 191–196.
viral liposome for gene therapy in cardiovascular diseases,
[98] C. Puyal, L. Maurin, G. Miquel, A. Bienvenue, J. Philippot,
Proc. Natl. Acad. Sci. U.S.A. 93 (1996) 11421–11425.
Design of a short membrane-destabilizing peptide covalently
Combination Therapy for Uncomplicated Falciparum Malaria in Ugandan Children A Randomized Trial Context Combination therapy is now widely advocated as first-line treatment for uncomplicated malaria in Africa. However, it is not clear which treatment regimensare optimal or how to best assess comparative efficacies in highly endemic areas. Objective To compare the efficacy and safety of 3
About myself I’m not a professional mathematician, I work as a pro-grammer. I have been studying in a university in Russia but have notfinished my study. So, I know little beyond my specialization. Nevertheless in my free time I discovered a new theory whichwould completely overturn general topology. About this lesson In this lesson I present my discovery, the theory of funcoidsand reloid
 Advanced Drug Delivery Reviews 30 (1998) 73–83
Recent advances in liposome technologies and their applications for systemic
aInex Pharmaceuticals Corporation, 1779 West 75th Street, Vancouver, BC, V6P 6P2, Canada
bDepartment of Biochemistry and Molecular Biology, The University of British Columbia, Vancouver V6T 1Z3, Canada
Received 1 July 1997; accepted 11 July 1997
Abstract
Advanced Drug Delivery Reviews 30 (1998) 73–83
Recent advances in liposome technologies and their applications for systemic
aInex Pharmaceuticals Corporation, 1779 West 75th Street, Vancouver, BC, V6P 6P2, Canada
bDepartment of Biochemistry and Molecular Biology, The University of British Columbia, Vancouver V6T 1Z3, Canada
Received 1 July 1997; accepted 11 July 1997
Abstract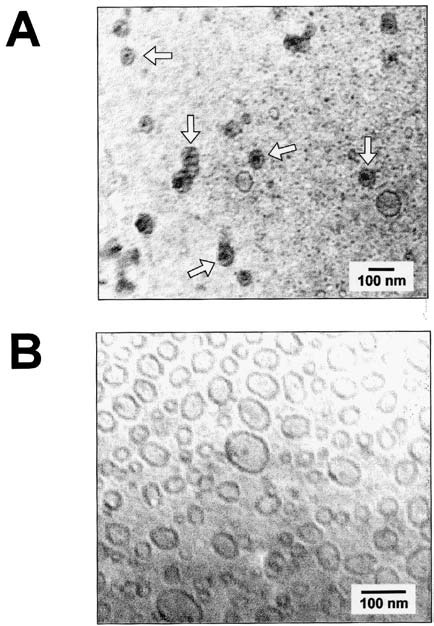 A. Chonn, P.R. Cullis / Advanced Drug Delivery Reviews 30 (1998) 73 –83
in transfecting KB cells in vitro, being 20–30 times
mid DNA–cationic lipid complex formed by the
more active than 3-b-[N-(N9,N9-dimethylethane)car-
addition of cationic lipids, added in monomer or
bamoyl]cholesterol–DOPE, (6:4, mol / mol) cationic
micellar form, to plasmid DNA [45,46]. This com-
liposome–plasmid DNA complexes. The stability
plex can serve as a well-defined intermediate in the
and pharmacokinetics of these particles upon in-
preparation of plasmid DNA-containing liposomes
travenous administration, or the ability of these
with good properties for systemic gene delivery
particles to transfect cells in vivo, have not been
applications (unpublished results). For example, the
Recently, Hofland et al. [42] described a detergen-
dioleoylphosphatidylcholine or DOPE, to these inter-
t-based procedure to form stable plasmid DNA–
mediates results in the formation of plasmid DNA-
lipidic particles by the addition of various amounts
containing liposomes that have a narrow size dis-
of cationic lipids [2,3-dioleoyloxy-N-(2(sperminecar-
tribution, averaging 70–100 nm in diameter (Fig. 1).
A. Chonn, P.R. Cullis / Advanced Drug Delivery Reviews 30 (1998) 73 –83
in transfecting KB cells in vitro, being 20–30 times
mid DNA–cationic lipid complex formed by the
more active than 3-b-[N-(N9,N9-dimethylethane)car-
addition of cationic lipids, added in monomer or
bamoyl]cholesterol–DOPE, (6:4, mol / mol) cationic
micellar form, to plasmid DNA [45,46]. This com-
liposome–plasmid DNA complexes. The stability
plex can serve as a well-defined intermediate in the
and pharmacokinetics of these particles upon in-
preparation of plasmid DNA-containing liposomes
travenous administration, or the ability of these
with good properties for systemic gene delivery
particles to transfect cells in vivo, have not been
applications (unpublished results). For example, the
Recently, Hofland et al. [42] described a detergen-
dioleoylphosphatidylcholine or DOPE, to these inter-
t-based procedure to form stable plasmid DNA–
mediates results in the formation of plasmid DNA-
lipidic particles by the addition of various amounts
containing liposomes that have a narrow size dis-
of cationic lipids [2,3-dioleoyloxy-N-(2(sperminecar-
tribution, averaging 70–100 nm in diameter (Fig. 1).