Tadalafil entfaltet seine Wirkung über eine selektive Hemmung der PDE5, wodurch die Konzentration von cGMP im glatten Muskelgewebe stabil bleibt. Diese biochemische Modulation resultiert in einer langanhaltenden Relaxation der Gefäßwände. Der Wirkstoff wird nach oraler Einnahme effizient resorbiert, mit einer Bioverfügbarkeit von rund 80 %. Seine Halbwertszeit von bis zu 36 Stunden ist innerhalb dieser Substanzklasse außergewöhnlich. Abgebaut wird er in der Leber, hauptsächlich durch CYP3A4, mit anschließender biliärer Exkretion. Typische unerwünschte Wirkungen entstehen durch eine verstärkte Vasodilatation, etwa Kopfschmerzen oder Flush. Pharmakologisch wird cialis generika vor allem durch die verlängerte Wirkungsdauer charakterisiert.
080869u165
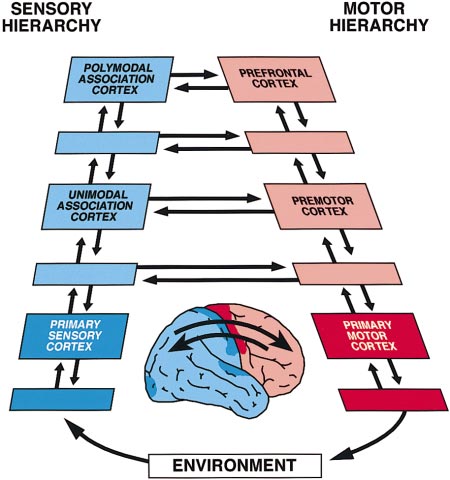
Neuron, Vol. 21, 1223–1229, December, 1998, Copyright 1998 by Cell Press Previews memory code essentially a relational code. According Linkage at the Top to this view, memory networks, after they have been formed, are defined by their cortico-cortical connectiv- ity, are exquisitely specific with regard to their content, Around the last turn of the century, neuroscientists were and presumably, to a degree, are topographically idio- heatedly debating the associative functions of the large syncratic for each individual. In the course of behavior, regions of the cerebral cortex that lie between primary reasoning, or speech, memory networks are succes- sensory and motor areas. Primary areas had recently sively activated—ignited, to use Braitenberg’s term been identified anatomically, and their functions were (1978)—by recall, recognition, or the need to retain them being unveiled by lesion and electrical methods. Little in short-term memory. Working memory, the kind of or nothing was known, however, about those other areas short-term memory needed to perform a sequential task, between them, which in the human included wide ex- may simply be the temporary activation of a widespread panses of the cortex of the occipital, temporal, and pari- cortical network of long-term memory for prospective etal lobes, as well as the prefrontal cortex, by itself action. That prospective action is what determines the making up nearly one-third of the neocortex. Flechsig role of the prefrontal cortex in that state of memory, for (1901), noting that those large areas developed late in this cortex contains the action-related associations of phylogeny and ontogeny, proposed that they served to networks originating in posterior cortex and itself plays mediate new associations of sensation with movement a critical role in the organization of sequential actions and also certain associative functions of the mind, such toward a goal (Fuster, 1997b). as memory. Despite respectable support from clinical To better understand the interactions between the observations and animal experiments, those ideas were prefrontal and other cortices in behavior, it is useful to dismissed, even ridiculed, as unfounded attempts to view them all in the context of the perception–action revitalize, by neurologizing it, a fading doctrine of associ- cycle. Briefly, this cycle is the circular flow of neural ationist psychology. For most of this century, the neuro- information by which an organism relates to its environ- physiologists of the cortex have ignored them. The thala- ment, a basic principle of biological cybernetics. The mus has been widely considered the key to cortical figure shows very schematically the cortical stages of physiology, and the sensory and motor cortices the key that cycle and their interconnections (human brain in to the physiology of the areas beyond. the inset, arrows symbolizing aggregates of fiber con- That began to change some 30 years ago. Since then, nections demonstrated in the monkey). Those cortical some of the old ideas have come back, this time bol- stages are the upper levels of two parallel hierarchies stered by solid anatomical and physiological evidence. of neural structures, one sensory and the other motor, We have discovered the previously unsuspected rich- that extend through the entire length of the nerve axis, ness and specificity of cortico-cortical axonal connec- from the spinal cord to the highest cortex of association. tions (Pandya and Yeterian, 1985). Even in primary visual (The unlabeled stages represent intermediary cortical cortex, which critically depends on the thalamus, Ͻ5% of the terminal axons have been found to be of thala- mic—geniculate—origin; the vast majority are of cortical origin, local or remote (Peters and Payne, 1993). High- lighting the importance of cortical connectivity for cogni- tive functions, microelectrode recording studies in the behaving monkey have revealed the widespread cortical activation of neurons during the memorization of an event or an object. We are rapidly making the transition from modular cognition to network cognition. The new perspective is that memory representations are com- prised of widely distributed cortical networks that tran- scend areas and modules by any anatomical definition (reviewed by Fuster, 1997a). Memory networks are prob- ably hierarchically organized, overlapping anatomically, and profusely interconnected. Accordingly, any neuron or group of neurons, anywhere in the cortex, can be part of many networks and thus many memories. The making of those networks follows certain princi- ples of synaptic modulation that are not yet fully under- stood. It appears almost certain, however, that learning and the acquisition of memory are based on the synaptic linkage of elementary cortical representations or nets into complex networks. Those new or expanded net- works represent new cognitive structures or gestalts. All memory would, therefore, be associative and the The Cortical Anatomy of the Perception–Action Cycle Neuron 1224 areas or subareas of adjacent labeled regions.) All con- and thus also to become engaged in the interactions at nections between stages are bidirectional, providing the summit of the perception–action cycle. In general, feedforward as well as feedback. however, as sensory–motor associations become rou- In the course of new or recently acquired behavior, tine, they are presumably relegated to lower stages of sensory information is processed along the sensory hi- the cycle. That is probably why, with overlearning, corti- erarchy—both serially and in parallel. In the cortex, that cal activations disappear from tomographic screens, information translates into action, which is processed and the neurons described by Asaad et al. seem to down the motor hierarchy to produce change in the lose their interest in old or familiar associations. The environment, which leads to sensory change, which is experimental approach of these investigators is uniquely processed through the sensory hierarchy and then mod- suited to reveal these changes. Indeed, somewhat para- ulates further action, and so on. The prefrontal and pos- doxically, the microelectrode remains the best tool to terior association cortices are in the cycle inasmuch and explore neural mechanisms in distributed cortical net- for as long as the behavior contains novelty, uncertainty, works with thousands if not millions of neurons. or ambiguity and has to bridge time spans with short- term memory. As those constraints disappear and be- Joaquı´n M. Fuster havior becomes automatic (e.g., walking, skilled rou- Neuropsychiatric Institute tines), the action is integrated in lower structures (e.g., University of California premotor cortex, basal ganglia) and sensory processing Los Angeles, California 90024 shunted at lower levels of the cycle. Asaad et al. (1998 [this issue of Neuron]) take us closer Selected Reading than ever before to understanding how those action- Braitenberg, V. (1978). In Theoretical Approaches to Complex Sys- related associations are formed in the prefrontal cortex, tems, R. Heim and G. Palm, eds. (Berlin: Springer), pp. 171–188. at the top of the cycle. Their experimental animal, a Flechsig, P. (1901). Lancet 2, 1027–1029. monkey, is trained in a delay task, where a particular Funahashi, S., Bruce, C.J., and Goldman-Rakic, P.S. (1989). J. Neu- visual stimulus calls for a particular movement of the rophysiol. 61, 331–349. eyes after a short delay. This delay makes the task a Fuster, J.M. (1997a). Trends Neurosci. 20, 451–459. memory task, requiring the subject to recognize and Fuster, J.M. (1997b). The Prefrontal Cortex (Philadelphia: Lippincott- retain a stimulus for subsequent action. Based on previ- ous research, so-called memory cells are expectedly Fuster, J.M., and Alexander, G.E. (1971). Science 173, 652–654. found, which fire faster during the delay than during Fuster, J.M., and Jervey, J. (1981). Science 212, 952–955. intertrial baseline periods; the discharge of some of Miller, E.K., Li, L., and Desimone, R. (1993). J. Neurosci. 13, 1460– these cells is stimulus preferential, that is, higher in reac- tion to a given stimulus than to another. In other cells Niki, H. (1974). Brain Res. 70, 346–349. nearby, the discharge is related to the movement. Most Pandya, D.N., and Yeterian, E.H. (1985). In Cerebral Cortex, A. Peters notable is the finding of cells that are related to both and E.G. Jones, eds. (New York: Plenum), pp. 3–61. the cue and the response, or a particular combination of Peters, A., and Payne, B.R. (1993). Cereb. Cortex 3, 69–78. the two. As the learning of a new association progresses, Zhou, Y., and Fuster, J.M. (1997). Exp. Brain Res. 116, 551–555. activity in prefrontal cells related to the direction of impending movement develops progressively earlier. Thus, the authors demonstrate in an elegant manner that prefrontal neurons become part of cortical networks containing and representing associations between vi- sual stimuli and movements. Touch Channels Sense Because memory cells were observed first in the pre- Blood Pressure frontal cortex and repeatedly reencountered in it (Fuster and Alexander, 1971; Niki, 1974; Funahashi et al., 1989), such cells have long been considered the substrate of its specific role in working memory. There is now ample Although we can all cite examples of individuals that evidence, however, that this state of memory activates seem to operate without perfusing their brains, this is also other broad and widely dispersed areas of the cor- just an illusion. Nature has installed pressure sensors tex with which the prefrontal cortex is connected. In (baroreceptors) to ensure relatively constant blood flow addition to prefrontal neurons, the short-term retention through their arteries. Imbedded in the walls of the arch of visual stimuli elicits the sustained activation of neu- of the aorta and the carotid sinus, arterial baroreceptor rons in inferotemporal cortex (Fuster and Jervey, 1981; nerve termini form intricate networks that fire in re- Miller et al., 1993) and even in somatosensory cortex if sponse to changes in blood pressure. These nerves the task is visuo-haptic (Zhou and Fuster, 1997). In sum, report to the brain stem respiratory centers located in therefore, the memory-active prefrontal cells are part of the solitary tract nucleus. In turn, these centers regulate extensive networks that span posterior as well as frontal blood vessel tone and heart pumping effectiveness cortex. There is evidence that their sustained activation through the sympathetic nervous system. In this issue in working memory results from the dynamic interac- of Neuron, Drummond et al. (1998) provide evidence tions between those cortices at or near the top of the that the mechanotransducers for the arterial pressure perception–action cycle (Fuster, 1997a). The cells that sensors are members of the degenerin (DEG)/ENaC fam- Asaad et al. describe seem to become part of those ily of cation channels. networks as they are formed or expanded by learning Although the baroreceptor reflex is well understood, Previews 1225 little is known about the basic mechanosensory process Mammalian homologs, BNaC1, BNaC2, and DRASIC, that senses distension of the arterial wall. Ion channels have also been cloned from nervous tissue (reviewed whose gating is responsive to changes in plasma mem- by Tvernarakis and Driscoll, 1997; Snyder et al., 1998, brane tension are primary candidates for these mecha- and references therein). Mutations near the second trans- notransducers. In the cardiovascular system, mechano- membrane domain result in the DEG/ENaC channels sensitive channels have been recorded from endothelial being constitutively open, allowing the unobstructed en- cells lining the lumen of arteries and from cardiac myo- try of cations into the cell. The flood of cations results cytes (reviewed by Sachs and Morris, 1998). The energy in degeneration of the mechanosensory neurons of C. needed to gate mechanosensitive channels may be col- elegans. lected by the membrane-associated cytoskeleton. But, Can the DEG/ENaC channels bridge the mechanosen- to date, the only cloned channel that is an unequivocal sory gap between arterial blood pressure and barore- mechanosensor is the bacterial MscL protein (reviewed ceptor discharge? Until recently, the evidence linking by Sukharev et al., 1997). The bacterial channel is unique baroreceptor mechanotransduction with the DEG/ENaC in that it is a hexameric protein complex that can be channels was purely circumstantial. Mechanosensitive gated by membrane tension independent of cytoskeletal gating of the DEG/ENaC channels has not been unequiv- elements. ocally shown in their native tissues, in part due to the Our first glimpse at the molecular structure of a mech- relative inaccessibility of the baroreceptive nerve termi- anosensitive channel in eukaryotes was obtained from nals buried within the arterial wall. Nonetheless, mecha- genetic studies conducted in the worm Caenorhabditis nosensory responses have been observed from dissoci- elegans. These worms move away in response to light ated baroreceptor neurons isolated from the nodose touch of the nose or body. Using genetic approaches, ganglion, which innervates the aortic arch. These re-
ف400 mutants were isolated that were defective in the sponses included macroscopic Ca2ϩ entry in cells in touch response but still capable of locomotion (reviewed response to membrane distortion by a puff of solution by Tvernarakis and Driscoll, 1997). From these mutants, (Sullivan et al., 1997) and single channel cation currents 16 genes were identified that when mutated gave rise activated by suction applied through a recording elec- to the aberrant mechanosensory phenotype, Mec. The trode (Kraske et al., 1998). Although these responses Mec mutations involve proteins localized in a network were blocked by the trivalent gadolinium previously shown of six neurons and associated cytoskeletal and extracel- to block mechanosensitive channels in other prepara- lular components. These proteins are distributed across tions, their sensitivity to amiloride was not demonstrated the long axis of the worm and comprise what are now (Hamill and McBride, 1996). Also, since the site of mech- known as touch receptors. Interestingly, mutations within anotransduction is at the nerve terminals imbedded in a subset of these genes also result in neuronal cell death the arterial wall, the significance of mechanosensitive and are hence also broadly referred to as degenerins responses measured on the soma is questionable. Drummond et al. (1998) use reverse transcriptase A subset of the DEG proteins (MEC-4, MEC-10) share polymerase chain reaction (RT–PCR) to show that  and homology with the amiloride-sensitive sodium channel
␥ subunits of the epithelial amiloride sodium channel subunits previously described in the epithelial layers of (ENaC) are present in isolated baroreceptor cells of the the kidneys, lungs, and intestines of vertebrates (Palmer, nodose ganglion. Since nodose ganglia contain non- 1992). The epithelial amiloride-sensitive sodium channel baroreceptor cells, this result was corroborated by im- (ENaC) is a multimeric protein complex composed of munostaining for ␥ENaC in baroreceptor neurons spe- three subunits (␣, , and ␥), each of which is thought cifically labeled with the fluorescent lipophilic dye Di-I. to be represented three times in the channel complex Di-I applied to the aortic arch retrogradely labeled a (Snyder et al., 1998). This finding inspired the notion that majority (80%) of nodose cells that had also stained MEC-4 and MEC-10 comprise subunits of a mechani- positively for ␥ENaC. Anterogradely labeled nodose cally gated ion channel related to the amiloride-sensitive ganglia stained small nerve terminals in the aortic arch epithelial sodium channel, and, indeed, amiloride is known with both Di-I and anti-␥ENaC, and the labeled nerve to block certain classes of mechanosensitive channels terminals had complicated morphologic features pre- (Hamill and McBride, 1996). As a family, these proteins viously associated with baroreceptor nerve terminals. have been termed the DEG/ENaC cation channels. Surprisingly, ␣ENaC subunit could not be demonstrated Structurally, each DEG/ENaC channel subunit con- in nodose ganglia, raising the possibility that ␥ and tains two hydrophobic transmembrane segments, a large
ENaC subunits might be associating with an unidenti- extracellular loop containing three cysteine-rich regions, fied third channel subunit. This result may underlie the a domain with homology to venom neurotoxins, and differences in mechanosensitive channel conductance cytoplasmic N and C termini through which the channel and selectivity previously observed in a variety of tissue is thought to associate with the cytoskeleton. Interest- types (Sachs and Morris, 1998). Finally, mechanosen- ingly, the bacterial MscL channel also contains two sory responses, such as puff-induced Ca2ϩ entry in membrane-spanning domains and cytoplasmic N and retrogradely labeled nodose cells and baroreflex nerve C termini. Other MEC proteins include tubulin-based cytoskeletal proteins (MEC-2, MEC-7, MEC-12) and com- discharge in response to artery distention, could be re- ponents of the extracellular matrix (MEC-5 and MEC-9). versibly inhibited by amiloride and its analog. Although DEG/ENaC homologs also exist in C. elegans but are not demonstrating mechanosensitive gating of DEG/ not confined to the touch receptor complexes. UNC-8 ENaC channels directly, these results do strengthen the and DEL-1 are DEG/ENaC homologs expressed in motor evidence that these channels are the basic mechano- neurons, while UNC-105 is expressed in muscle cells. transducers in the baroreceptor nerve terminals. Neuron 1226 What is the role of the membrane-associated cyto- brain are likely to be complex, and the tools we posses skeleton in gating the DEG/ENaC channels? The Unc- are relatively coarse. In this light, the fact that scientists 105 mutant in C. elegans is characterized by hypercon- generally are clever enough to think of mechanistic sce- tracted muscle resulting from unabated cation entry. narios that cannot be disproved by existing empirical Unc-105 interacts with Let-2, which encodes collagen tools complicates the search. Furthermore, the imbal- IVa2. The Unc-105 mutation can be counteracted by anced impact of positive results over negative results, mutations in Let-2, further reinforcing the notion that or the natural bias of scientists to champion their own the cytoskeleton is important in the gating of mechano- point of view, can prolong the discourse. Whatever the sensitive channels. In humans, X-linked Becker’s and source, the field of LTP has been mired with LTC to the Duchenne’s muscular dystrophies are associated with point that most consider it a long-term tar pit (LTTP). a faulty myoplasmic Ca2ϩ handling somehow resulting How does one escape eternal fossilization? It can only from the disruption of the cell cytoskeleton (Anderson be hoped that over time different groups, using different and Kunkel, 1992). By analogy, recordings of mechano- techniques and asking questions related to different as- sensitive channels from skeletal muscle from a mouse pects of synaptic transmission modulation, will provide model of human X-linked muscular dystrophy (mdx) ex- the cleansing solvent. hibit constitutively active channels at rest (Franco and Toward this end, a number of groups have been Lansman, 1990) and elevated Ca2ϩ entry (Turner et al., scouring the biophysical underpinnings of some scenar- 1991). It will be interesting to see if mutations of the DEG/ ios proposed to explain LTP in CA1 hippocampus. This ENaC channels cause human disorders not previously month, Gomperts et al. (1998 [this issue of Neuron]) understood on the molecular level. address the biophysical basis of “silent” synapses, a sticky issue currently at the fulcrum of the debate over Alfredo Franco-Obrego´n and David E. Clapham whether LTP is due to a pre- or postsynaptic modifica- Children’s Hospital tion. “Silent” synapses refer to excitatory transmission Harvard Medical School mediated purely by NMDA receptors (NMDARs): due Department of Basic Cardiovascular Research to the voltage-dependent properties of NMDARs, such Boston, Massachusetts 02115 transmission will produce no postsynaptic response at resting potentials; hence, it is termed silent. Addition of Selected Reading AMPARs (which are functional at resting potentials) to synapses with only NMDARs was proposed as a possi- Anderson, M.S., and Kunkel, L.M. (1992). Trends Biochem. Sci. 17, ble postsynaptic mechanism to explain the (consistently 289–292. observed) decrease in synaptic failures during LTP, evi- Drummond, H.A., Price, M.P., Welsh, M.J., and Abboud, F.M. (1998). Neuron 21, this issue, 1435–1441. dence that is traditionally interpreted as a presynaptic Franco, A., and Lansman, J.B. (1990). Nature 344, 670–673. change (Liao et al., 1992). Support for such a process, relying on the difference in variability between AMPAR- Hamill, O.P., and McBride, D.W. (1996). Pharmacol. Rev. 48, 231–252. and NMDAR-mediated responses, was initially detected Kraske, S., Cunningham, J.T., Hajduczok, G., Chapleau, M.W., Ab- by Kullmann (1994). This view was strengthened by di- boud, F.M., and Wachtel, R.E. (1998). Am. J. Physiol. 275, H1497– rect observations of pure NMDAR-mediated synaptic responses and a conversion of silent synapses to func- Palmer, L.G. (1992). Annu. Rev. Physiol. 54, 51–66. tional synapses during LTP (Isaac et al., 1995; Liao et Travernarakis, N., and Driscoll, M. (1997). Annu. Rev. Physiol. 59, al., 1995). Thus, a simple postsynaptic model emerged 659–689. that could largely explain the existing data on LTP, even Turner, P.R., Fong, P.Y., Denetclaw, W.F., and Steinhardt, R.A. those data classically interpreted as a change in presyn- (1991). J. Cell Biol. 115, 1701–1712. aptic function. If nothing else, this model is attractive Sachs, F., and Morris, C.E. (1998). Rev. Physiol. Biochem. Pharma- because it requires only established intracellular sig- cology 132, 1–77. naling mechanisms. It has been well accepted that post- Snyder, P.M., Cheng, C., Prince, L.S., Rogers, J.C., and Welsh, M.J. synaptic processes initiate LTP; now well-established (1998). J. Biol. Chem. 273, 681–684. intracellular second messenger mechanisms (such as Sukharev, S.I., Blount, P., Martinac, B., and Kung, C. (1997). Annu. protein phosphorylation or membrane trafficking) can Rev. Physiol. 59, 633–657. explain the longer-lasting modification. Sullivan, M.J., Sharma, R.V., Wachtel, R.E., Chapleau, M.W., Waite, L.J., Bhalla, R.C., and Abboud, F.M. (1997). Circ. Res. 80, 861–867. However, this model requires the existence of syn- apses with only NMDARs. While few doubt that pure NMDAR responses exist, an alternative mechanism to the silent synapse hypothesis has been proposed based on a series of experimental findings (reviewed by Kull- mann and Asztely, 1998). In this scenario, all excitatory Silencing the Controversy in LTP? synapses have both AMPA and NMDA receptors. Pure NMDA responses onto cell A are due to the “spillover” of transmitter from a synapse directly contacting cell B. Why has there been such long-term controversy (LTC) The concentration of transmitter, once it reaches cell over the mechanisms underlying long-term potentiation A, is sufficient to activate NMDARs but not AMPARs (LTP)? The inability to resolve this debate may have because of their lower affinity for transmitter. Gomperts many sources, including intrinsically empirical as well et al. test this model by examining excitatory transmis- as sociological factors. Certainly, the regulatory mecha- sion in a preparation where an individual neuron is cul- nisms underlying modification of transmission in the tured in isolation and makes synapses only on itself. In Previews 1227 Selected Reading this case, every presynapse in the preparation forms a direct contact on the recorded neuron. If every synapse Diamond, J.S., Bergles, D.E., and Jahr, C.E. (1998). Neuron 21, has both AMPA and NMDA receptors, even a spillover 425–433. response will always be accompanied by a direct re- Gomperts, S.N., Rao, A., Craig, A.M., Malenka, R.C., and Nicoll, R.A. sponse, i.e., a response with an AMPA component. (1998). Neuron 21, this issue, 1443–1451. Gomperts et al. detect pure NMDA responses in this Isaac, J.T., Nicoll, R.A., and Malenka, R.C. (1995). Neuron 15, preparation in several ways. First, they note that evoked 427–434. responses have a larger NMDAR component than spon- Kullmann, D.M. (1994). Neuron 12, 1111–1120. taneous miniature responses that are selected based Kullmann, D.M., and Asztely, F. (1998). Trends Neurosci. 21, 8–14. on having an AMPAR component. This suggests that Liao, D., Jones, A., and Malinow, R. (1992). Neuron 9, 1089–1097. a significant number of spontaneous events were not Liao, D., Hessler, N.A., and Malinow, R. (1995). Nature 375, 400–404. selected that have an NMDAR component and no Liao, D., Zhang, X., O’Brien, R., Ehlers, M., and Huganir, R.L. (1999). AMPAR component. Furthermore, they are able to pick Nat. Neurosci., in press. out spontaneous events that, when averaged, have a Lu¨scher, C., Malenka, R.C., and Nicoll, R.A. (1998). Neuron 21, 435–441. slow time course similar to that of a pure NMDAR re- Mainen, Z.F., Jia, Z., Roder, J., and Malinow, R. (1998). Nat. Neurosci. sponse. These results strongly suggest that pure NMDA 7, 579–586. responses can be detected in this preparation and thus Nusser, Z., Lujan, R., Laube, G., Roberts, J.D., Molnar, E., Somogyi, argue that there must be some mechanism, other than P. (1998). Neuron 21, 545–559. spillover, to account for pure NMDA responses. Petralia, R.S., Esteban, J.A., Wang, Y.-X., Partridge, J.G., Zhao, Gomperts et al. examine this further: using immuno- H.-M., Wenthold, R.J., and Malinow, R. (1999). Nat. Neurosci., in labeling techniques, they make two important observa- tions. First, all presynaptic boutons have a cluster of adjacent postsynaptic receptors. Thus, indeed, any spillover response would also produce a direct re- sponse. Secondly, they show that a significant fraction of synaptic connections have NMDA and lack AMPA Eph Receptors, Ephrins, and PDZs receptor immunolabeling and can thus account for the pure NMDAR transmission. Gather in Neuronal Synapses Thus, for this preparation, the authors argue that trans- mitter spillover cannot account for the pure NMDAR responses, and they provide anatomical evidence for Efficient intercellular communication depends on the synapses with only NMDARs. This, along with another localization of specific signaling proteins to particular recent study (Liao et al., 1999), indicates that cultured sites on the cell surface. The synaptic junction, which neuronal preparations have silent synapses that can be mediates rapid communication between neurons, pro- accounted for by synapses with only NMDARs. Such vides a striking example in which specific proteins ac- synapses have also been identified in the experimentally cumulate at membrane specializations on both sides of more hostile terrain of the intact brain with immunogold the synapse. For instance, ionotropic glutamate re- electron microscopy (Nusser et al., 1998; Petralia et al., ceptors are highly concentrated in the postsynaptic 1999). Pure NMDAR synapses were found to be more membrane of excitatory synapses. What is the molecu- prevalent in CA1 hippocampus early in postnatal devel- lar mechanism underlying such localized clustering of opment, supporting the view that initial synapses my membrane proteins? Recent studies have highlighted the role played by proteins that contain PDZ domains be silent and become AMPAfied during development (Sheng, 1997; Ziff, 1997). PDZ domains are modular pro- through an activity-dependent process (Nusser et al., tein interaction domains that typically recognize short peptide sequences of four or more amino acids at the Finding that silent transmission can be due to action at very C terminus of its ligands, and different PDZ domains synapses with only NMDARs enhances our knowledge recognize different C-terminal sequences. For example, about basic excitatory transmission in the brain. Further- PDZ domains in the PSD-95/SAP90 family of postsynap- more, this provides an important element to a postsyn- tic density proteins bind to the C-terminal -ESDV peptide aptic model for expression of LTP. These results come at sequence of NR2 subunits of the NMDA receptor. On the heel of several studies arguing against presynaptic the other hand, GluR2/3 subunits of AMPA receptors changes during LTP. Three independent groups, using bind via their C termini (-SVKI) to GRIP, a protein con- synaptic (Mainen et al., 1998) or peri-synaptic (Diamond taining seven PDZs (Dong et al., 1997). Studies of PDZ- et al., 1998; Lu¨scher et al., 1998) detectors of synaptic based interactions in synapses have naturally focused transmitter release, found no increase after LTP. While on neurotransmitter receptors and ion channels, which an optimistic observer may thus conclude that the tar are known to be concentrated in synaptic junctions. By is thinning, and that the LTC of LTP is getting resolved, contrast, little is known about receptor tyrosine kinases there may (always) be more clever scenarios to consider. (RTKs) in neuronal synapses. Some RTKs (MuSK and erbB receptors) are concentrated in the vertebrate neu- romuscular junction, but the mechanisms underlying Roberto Malinow this localization are unclear. No interactions between Cold Spring Harbor Laboratory RTKs and PDZ domains have been reported in verte- Cold Spring Harbor, New York 11724 brates. Enter Torres et al. (1998 [this issue of Neuron]) Neuron 1228 with two significant advances. First, they report that specific protein complex around their membrane protein RTKs of the Eph family and their transmembrane ligands ligands. In Drosophila photoreceptors, a physiologically (ephrins) bind to specific PDZ domain proteins; second, coupled “transducisome” of phototransduction signal- certain Eph receptors and ligands are concentrated in ing proteins is built around InaD, a protein with five neuronal synapses, probably in association with their PDZs (Tsunoda et al., 1997). In synapses, PSD-95 can PDZ binding partners. assemble a specific cytoskeletal-signaling complex that Torres et al. show that the Eph RTK EphB2 (C-terminal is physically linked to the NMDA receptor (Craven and sequence -SVEV) has specific affinity for PDZ domains Bredt, 1998). Perhaps the interaction of Eph receptors in two different proteins, GRIP and PICK1, while EphA7 and ligands with PDZ proteins couples them to intracel- (-GIQV) can bind to GRIP, PICK1, and a third PDZ-con- lular signaling networks or modulatory enzymes. This taining protein, syntenin. Ligands belonging to the may be particularly significant for the ephrin-B ligands, ephrin-B subfamily (-YYKV) also bind to GRIP, PICK1, which participate in reciprocal signaling with their Eph and syntenin. The interaction between PDZ proteins and receptors despite lacking a catalytic domain. PICK1 has Eph receptors/ligands is not so surprising; after all, PDZ only one PDZ domain but was previously identified as domains recognize just the last few amino acids of their a protein kinase C (PKC)-binding protein (Staudinger et ligands, and this C-terminal “zipcode” can be appended al., 1997); thus, PICK1 could mediate the association of onto any class of protein. Indeed, the precedent for an PKC with specific Eph receptors and ligands. PICK1 interaction between an RTK (LET23) and a PDZ protein also appears to be a direct substrate for the Eph RTK (LIN-7) has been established in C. elegans epithelial cells (Torres et al., 1998). GRIP has seven PDZs and the po- (Kaech et al., 1998). More unexpected is the ensuing tential to scaffold an elaborate protein architecture finding that EphB2 and its ligand, Ephrin-B, are concen- around Eph receptors and their ligands. Since GRIP trated at synapses in cultured neurons, where their PDZ was originally identified as an AMPA receptor–binding partners GRIP and PICK1 are also localized. protein, it will be interesting to determine whether Eph To date, Eph receptors and their ligands have been receptors or ligands are physically and functionally cou- studied primarily in a developmental context. In the ner- pled to AMPA receptors in synapses. To date, there has vous system, these molecules are implicated in axon been little evidence for regulation of AMPA receptors guidance, particularly in repulsion and in establishment by tyrosine phosphorylation. of boundaries between groups of cells. What are Eph Unlike many ligands of RTKs, Eph ligands are not receptors and ephrins doing in synapses? It is tempting active as soluble proteins; ephrins need to be clustered to speculate that they might be involved in synaptogene- on the cell surface for them to stimulate their cognate sis (like MuSK and erbB receptors) or in synaptic plastic- Eph receptors. It is pertinent, therefore, that surface ity, perhaps by controlling the adhesion and/or repulsion aggregation of transmembrane proteins is a common of pre- and postsynaptic membranes. The synaptic lo- outcome of interaction with PDZ proteins. The ability of calization of Eph receptors and their ligands needs to certain PDZ proteins to cluster their binding partners be confirmed in the brain and extended to other mem- may reflect the propensity of PDZ-containing proteins to bers of these protein families. Important questions will multimerize and/or their ability to bind these membrane include whether the various Eph receptors and ephrins proteins in a multivalent manner. Indeed, PICK1 can are differentially distributed among CNS synapses, and aggregate ephrin-B1 in heterologous cells (Torres et al., whether receptors and ligands are segregated to pre- 1998). Clustering by PICK1 or GRIP may optimize the and postsynaptic sides of the junction. Detailed analysis presentation of ephrins to their Eph receptors in vivo; of mouse knockouts of Eph receptors and ephrins may such a mechanism offers another potential level for reg- shed more light on the roles of these proteins in syn- ulation of Eph signaling. PDZ-dependent clustering of apses and in mature brain. Eph receptors and ligands at specific subcellular sites If it is early to speculate about the synaptic functions (e.g., in growth cones) may also be important for Eph/ of Eph receptors and ephrins, what about the functional ephrin function in development. Thus, following up the significance of their interactions with PDZ domain pro- findings of Torres et al. promises to shed new light on teins? A prevailing idea is that the PDZ protein is impor- the functions and mechanisms of the Eph system in tant for the subcellular localization of its binding part- both developing and mature brain. ners. In Drosophila, the PSD-95 homolog Discs-large is localized in synapses and is essential for the synaptic Yi-Ping Hsueh and Morgan Sheng clustering of its PDZ interactors, Shaker and Fasciclin Howard Hughes Medical Institute and Department II (Thomas et al., 1997; Zito et al., 1997). Genetic studies of Neurobiology on InaD (in Drosophila) and LIN-2/LIN-7/LIN-10 (in C. Massachusetts General Hospital and elegans) additionally support the idea that PDZ-medi- Harvard Medical School ated interactions are important for the subcellular tar- Boston, Masachusetts 02114 geting of the interacting proteins, both at synapses and at other specialized membrane domains (Tsunoda et al., 1997; Kaech et al., 1998; Rongo et al., 1998). By analogy, Selected Reading EphB2 and ephrin-B1 localization in neuronal synapses may depend on their binding to synaptic PDZ proteins Craven, S.E., and Bredt, D.S. (1998). Cell 93, 495–498. like GRIP and PICK1. Dong, H., O’Brien, R.J., Fung, E.T., Lanahan, A.A., Worley, P.F., and Another (not mutually exclusive) concept is that PDZ Huganir, R.L. (1997). Nature 386, 279–284. proteins have a scaffolding function and can assemble a Kaech, S.M., Whitfield, C.W., and Kim, S.K. (1998). Cell 94, 761–771. Previews 1229 Rongo, C., Whitfield, C.W., Rodal, A., Kim, S.K., and Kaplan, J.M. (1998). Cell 94, 751–759. Sheng, M. (1997). Nature 386, 221–223. Staudinger, J., and Olsen, E.N. (1997). J. Biol. Chem. 272, 32019– 32024. Thomas, U., Kim, E., Kuhlendahl, S., Ho Koh, Y., Gundelfinger, E.D., Sheng, M., Garner, C.C., and Budnik, V. (1997). Neuron 19, 787–799. Torres, R., Firesrein, B.L., Dong, H., Staudinger, J., Olsen, E.N., Huganir, R.L., Bredt, D.S., Gale, N.W., and Yancopoulos, G.D. (1998). Neuron 21, this issue, 1453–1463. Tsunoda, S., Sierralta, J., Sun, Y., Bodner, R., Suzuki, E., Becker, A., Socolich, M., and Zuker, C.S. (1997). Nature 388, 243–249. Ziff, E.B. (1997). Neuron 19, 1163–1174. Zito, K., Fetter, R.D., Goodman, C.S., and Isacoff, E.Y. (1997). Neuron 19, 1007–1016.
Ear drops April 2013 Many divers complain about pain in their auditory canals during their diving holiday and have no idea how to deal with it. Every diver should therefore know something about this topic, so that a diving trip does not have to come to a premature and painful end. Some divers regularly get painful inflammations of their auditory canals, especial y when diving in war
PATIENT'S MEDICAL HISTORY PATIENTS NAME_____________________________________ DATE OF BIRTH_________________ ALTHOUGH DENTAL PERSONNEL PRIMARILY TREAT THE AREA IN AND AROUND YOUR MOUTH, YOUR MOUTH IS A PART OF YOUR ENTIRE BODY, HEALTH PROBLEMS THAT YOU MAY HAVE, OR MEDICATION THAT YOU MAY BE TAKING, COULD HAVE AN IMPORTANT INTERRELATIONSHIP WITH THE DENTISTRY THAT YOU WILL BE RECEIVING. THANK YOU F
 Neuron, Vol. 21, 1223–1229, December, 1998, Copyright 1998 by Cell Press
Neuron, Vol. 21, 1223–1229, December, 1998, Copyright 1998 by Cell Press