Tadalafil entfaltet seine Wirkung über eine selektive Hemmung der PDE5, wodurch die Konzentration von cGMP im glatten Muskelgewebe stabil bleibt. Diese biochemische Modulation resultiert in einer langanhaltenden Relaxation der Gefäßwände. Der Wirkstoff wird nach oraler Einnahme effizient resorbiert, mit einer Bioverfügbarkeit von rund 80 %. Seine Halbwertszeit von bis zu 36 Stunden ist innerhalb dieser Substanzklasse außergewöhnlich. Abgebaut wird er in der Leber, hauptsächlich durch CYP3A4, mit anschließender biliärer Exkretion. Typische unerwünschte Wirkungen entstehen durch eine verstärkte Vasodilatation, etwa Kopfschmerzen oder Flush. Pharmakologisch wird cialis generika vor allem durch die verlängerte Wirkungsdauer charakterisiert.
Molecularpsychiatry.uk-wuerzburg.de
The Journal of Neuroscience, March 15, 2001, 21(6):2178–2185
Functional Consequences of 5-HT Transporter Gene Disruption on 5-HT Receptor-Mediated Regulation of Dorsal Raphe and Hippocampal Cell Activity Clotilde Mannoury la Cour,1 Claudette Boni,1 Naı¨ma Hanoun,1 Klaus-Peter Lesch,2 Michel Hamon,1 and Laurence Lanfumey1
1Institut National de la Sante´ et de la Recherche Me´dicale U288, Neuropsychopharmacologie Mole´culaire, Cellulaire etFonctionnelle, Faculte´ de Me´decine Pitie´-Salpeˆtrie`re, 75634 Paris Cedex 13, France, and 2Department of Psychiatry,University of Wu¨rzburg, 97080 Wu¨rzburg, Germany
The consequences of the absence of 5-HT reuptake on the
trasted with those obtained with hippocampal slices in which
5-carboxamidotryptamine was equipotent to hyperpolarize
dorsal raphe nucleus and the hippocampus of knock-out mice
CA1 pyramidal neurons in both mutant and wild-type mice. As
lacking the serotonin transporter (5-HTT). Extracellular record-
expected from their mediation through 5-HT
ings showed that application of selective 5-HT reuptake inhib-
effects of ipsapirone and 5-carboxamidotryptamine were com-
itors such as paroxetine and citalopram onto brainstem slices
petitively inhibited by the selective 5-HT
resulted in a concentration-dependent inhibition of 5-HT neu-
100635 in both groups. These data showed that 5-HTT gene
ron firing in the dorsal raphe nucleus of wild-type 5-HTTϩ/ϩ
knock-out induced a marked desensitization of 5-HT
mice, but not 5-HTTϪ/Ϫ mutants. By contrast, the 5-HT
ceptors in the dorsal raphe nucleus without altering postsyn-
receptor agonists ipsapirone and 5-carboxamidotryptamine in-
receptor functioning in the hippocampus. Similar-
hibited the discharge in both groups. However, the potency
ities between these changes and those evoked by chronic
of these agonists was markedly decreased (by ϳ55- and ϳ6-
treatment with 5-HT reuptake inhibitors emphasize the exis-
fold, respectively) in 5-HTTϪ/Ϫ compared with 5-HTTϩ/ϩ an-
imals. Similarly, intracellular recordings showed that the po-
tency of 5-carboxamidotryptamine to hyperpolarize 5-HT
Key words: 5-HT transporter knock-out mice; 5-HT
neurons in the dorsal raphe nucleus was significantly lower
tors; dorsal raphe nucleus; hippocampus; desensitization; in
in 5-HTTϪ/Ϫ than in 5-HTTϩ/ϩ animals. These data con-
The involvement of the serotoninergic system in major psychiatric
1996) allowed the generation of an animal model with targeted
diseases, in particular mood disorders such as depression, is a well
disruption of this gene by homologous recombination (Bengel et
established clinical feature (Asberg et al., 1976; Cryan and Leo-
al., 1998). Indeed, the deletion of exon 2 results in an inactive
nard, 2000). Accordingly, to date, the most frequently used anti-
gene and the complete absence of 5-HT reuptake activity in the
depressants are the selective serotonin reuptake inhibitors
homozygous 5-HTTϪ/Ϫ mice. No apparent developmental alter-
(SSRIs), which act on the Naϩ/ClϪ-dependent 5-HT transporter
ations were noted in the null mutant mice, suggesting that major
(5-HTT) (Graham et al., 1989; Lesch, 1997). In the CNS, 5-HTT
compensatory mechanisms occur in these animals during embry-
seems to be essentially localized on serotoninergic neurons, at the
onic and subsequent neurodevelopment (Bengel et al., 1998).
level of somas, dendrites, axons, and terminals (Hensler et al.,
However, marked depletions of 5-HT and of its metabolite
1994; Sur et al., 1996; Tao-Cheng and Zhou, 1999), and only a
5-hydroxyindoleacetic acid in brain evidenced that adaptive
minor glial expression of this protein has also been reported by
changes in 5-HT neurotransmission do occur in 5-HTTϪ/Ϫ mu-
some authors (Hirst et al., 1998; Pickel and Chan, 1999). In any
tants (Bengel et al., 1998; Li et al., 1999; Fabre et al., 2000).
case, it is well established that the 5-HTT is responsible for the
Previous studies using biochemical and neuroendocrinological
primary mechanism of 5-HT inactivation in the CNS (Lesch,
approaches further investigated adaptive changes in 5-HT neu-
rotransmission in 5-HTTϪ/Ϫ mutants with particular attention to
Elucidation of the murine 5-HTT gene sequence (Chang et al.,
5-HT receptors. Indirect evidence of desensitization and down-
regulation of 5-HT1A autoreceptors in the dorsal raphe nucleus
Received Aug. 7, 2000; revised Dec. 22, 2000; accepted Jan. 4, 2001.
(DRN) (Li et al., 1999; Fabre et al., 2000) and 5-HT2A receptors
This research was supported by the Institut National de la Sante´ et de la
in the striatum and cerebral cortex (Rioux et al., 1999) has thus
Recherche Me´dicale and Bristol-Myers Squibb Foundation (Unrestricted Biomed-
ical Research Grant Program). C.M.C. was a recipient of a Fondation pour la
been reported in knock-out mice. Interestingly, similar changes in
Recherche Me´dicale fellowship during performance of this work. We are grateful to
these receptors have previously been shown to occur after chronic
pharmaceutical companies (Lundbeck, Pierre Fabre, SmithKline Beecham,
Troponwerke-Bayer, and Wyeth-Ayerst) for generous gifts of drugs.
blockade of 5-HT reuptake by SSRI (Chaput et al., 1986; Sanders-
Correspondence should be addressed to C. Mannoury la Cour, Institut National de
Bush et al., 1989; Jolas et al., 1994; Kreiss and Lucki, 1995; Le
la Sante´ et de la Recherche Me´dicale U 288, Neuropsychopharmacologie Mole´culaire,
Poul et al., 1995, 2000), thereby suggesting that the 5-HTTϪ/Ϫ
Cellulaire et Fonctionnelle, Faculte´ de Me´decine Pitie´-Salpeˆtrie`re 91, Boulevard de
l’Hoˆpital, 75634 Paris Cedex 13, France. E-mail: mannoury@idf.ext.jussieu.fr.
mutant mouse can be considered as a model of whole-life treat-
Copyright 2001 Society for Neuroscience 0270-6474/01/212178-08$15.00/0
ment with these drugs. Interestingly, Le Poul et al. (2000) recently
Receptors in 5-HT Transporter Knock-Out Mice
J. Neurosci., March 15, 2001, 21(6):2178–2185 2179
DRN and pyramidal cells in the CA1 area of the hippocampus were
recorded in current-clamp mode with 3 M KCl-filled electrodes (50 – 80
M⍀), while brain slices were superfused with ACSF (Corradetti et al.,
1998). Electrical signals were amplified with an Axoclamp 2A (Axon
These data led us to investigate further the functional status of
Instruments, Foster City, CA) and displayed on an oscilloscope and a
5-HT1A receptors in these two areas in 5-HTTϪ/Ϫ mutants
chart recorder. Traces were stored in a digital tape recorder (DTR 1202;
versus wild-type mice. For this purpose, both extracellular and
BioLogic; 48 kHz sampling frequency) and a computer using pClamp6
intracellular electrophysiological recordings of 5-HT
software (3–10 kHz sampling frequency; Axon Instruments) for off-line
measurements. Only neurons with stable resting membrane potential
ing neurons in brain slices were used to quantitatively assess their
(range, Ϫ50 to Ϫ90 mV) and input resistance (R
CA1 neurons and 100 –500 M⍀ for DRN cells) throughout the recording
session were included in the analysis. Membrane potential in response to
hyperpolarizing and depolarizing current pulses of 50/100 pA increments
MATERIALS AND METHODS
(range, Ϫ900 to ϩ500 pA) was measured before, during, and after tissue
superfusion with drugs added to the ACSF. To draw concentration–
Experiments were performed using homozygous 5-HTTϪ/Ϫ, heterozy-
response curves for the 5-HT1 agonist 5-carboxamidotryptamine (5-CT),
gous 5-HTTϩ/Ϫ, and wild-type 5-HTTϩ/ϩ littermates born from het-
the membrane potential was recorded while slices were superfused
erozygous mutants of C57BL6 genetic background. Genotyping was
with increasing concentrations of this ligand. Preliminary experiments
performed as described by Bengel et al. (1998). Animals were used at 2
(data not shown) demonstrated that for a given cell, consecutive appli-
months of age when their body weight in each genotype equally ranged
cations of increasing concentrations of 5-CT produced cumulative
between 20 and 25 gm. After weaning and sexing, males and females
concentration-dependent responses with a maximal effect equal to that
were housed separately in groups of six to eight animals per cage and
obtained with application of a single saturating concentration. Nonlinear
maintained under standard laboratory conditions (22 Ϯ 1°C; 60% rela-
regression fitting was performed using Prism 2.0 (GraphPad) software
tive humidity; 12 hr light/dark cycle; food and water available ad libitum).
facilities for the determination of concentration-dependent hyperpolar-
In addition, some experiments were performed using CD1, C57BL6, and
ization and decrease in Rin caused by 5-CT.
c129 control mice provided by the Centre d’Elevage R. Janvier (Le
Genest-St. Isle, France) and IFFA Credo (Lyon, France), respectively.
Procedures involving animals and their care were conducted in con-
All data are given as means Ϯ SEM. Extracellular and intracellular
formity with the institutional guidelines that are in compliance with
recording data were analyzed by one-way ANOVA and, in case of
national and international laws and policies (council directive number
significance ( p Ͻ 0.05), the F test for significant treatment effects was
87– 848, 19 October 1987, Ministe`re de l’Agriculture et de la Foreˆt,
followed by the two-tailed Student’s t test to compare the experimental
Service Ve´te´rinaire de la Sante´ et de la Protection Animale, permissions
groups with their controls. A value of p Ͻ 0.05 was considered to be
Preparation of slices of DRN and dorsal hippocampus. Mice were decap-
The following drugs were used: ipsapirone (Bayer-Troponwerke, Cologne,
itated, and the brains were rapidly removed and immersed in an ice-cold
Germany), 5-CT (Research Biochemicals, Natick, MA), paroxetine (Smith-
Krebs’ solution, bubbled continuously with an O
Kline Beecham, Harlow, UK), citalopram (Lundbeck, Copenhagen,
(95:5%). A block of tissue containing the DRN or the dorsal hippocam-
N-[2-[4-(2-methoxyphenyl)-1-piperazinyl]ethyl]-N-
pus was cut into sections (350- to 400-m-thick) in the same ice-cold
(2-pyridinyl)cyclohexane carboxamide (WAY 100635; Wyeth-Ayerst,
Krebs’ solution using a vibratome (Corradetti et al., 1998). Brainstem or
hippocampus slices were then immediately incubated at room tempera-
ture (20 –23°C) for at least 1 hr in an artificial CSF (ACSF) of the
following composition (mM): NaCl 126, KCl 3.5, NaH2PO4 1.2, MgCl2
In both the DRN and the hippocampus, electrophysiological re-
2 2, NaHC O3 25, D-glucose 11, maintained at pH 7.3 by
continuous bubbling with O2–CO2 mixture. A slice of either the DRN or
cordings under the various pharmacological conditions tested did
the CA1 hippocampal area was then placed on a nylon mesh, completely
not reveal any significant differences between males and females of
submerged in a small chamber, and superfused continuously with oxy-
the homozygous 5-HTTϪ/Ϫ, heterozygous 5-HTTϩ/Ϫ, or wild-
genated ACSF (34°C) at a constant flow rate of 2–3 ml/min (Corradetti
type phenotype. Accordingly, both males and females were used
Extracellular recordings of serotoninergic neurons in the dorsal raphe
indifferently in the experiments reported herein. nucleus. Extracellular recordings were made with glass microelectrodes
filled with 2 M NaCl (10 –15 M⍀). Cells were identified as 5-HT neurons
Extracellular recordings of DRN 5-HT neurons
according to the following criteria: biphasic action potentials and slow
and regular pattern of discharge (1.5–2.5 spikes/sec) (Trulson and Fred-
Because generation of the 5-HTTϪ/Ϫ knock-out model required
erickson, 1987; Jacobs and Azmitia, 1992). Firing was evoked in the
otherwise silent neurons by adding the ␣
the use of three different strains of mice (c129, CD1, and C57BL6)
ephrine (3 M) to the superfusing ACSF (VanderMaelen and Aghaja-
(Bengel et al., 1998), some heterogeneity in the genetic background
nian, 1983). Baseline activity was recorded for 5–10 min before the
might have still existed, thereby accounting for possible variations
application of drugs via a three-way tap system that allowed complete
in the electrophysiological characteristics of DRN 5-HT neurons in
exchange of fluids within 2 min of arrival of a new solution. The electrical
signals were fed into a high-input impedance amplifier (VF180; Bio-
5-HTϪ/Ϫ mutants compared with wild-type animals of these
Logic, Claise, France), an oscilloscope, and an electronic ratemeter
strains. To directly assess this possibility, the spontaneous dis-
triggered by individual action potentials connected to an analog-to-
charge frequency of DRN 5-HT neurons was compared in paired
digital converter and a personal computer (Haj-Dahmane et al., 1991).
control 5-HTTϩ/ϩ mice and in c129, CD1, and C57BL6 mice.
The integrated firing rate was computed and recorded graphically as
Indeed, the baseline firing rate of 5-HT neurons was similar in the
consecutive 10 sec samples. The effect of a given drug was evaluated by
comparing the mean discharge frequency during the 2 min before its
four groups: c129, 1.72 Ϯ 0.19 spikes/sec (mean Ϯ SEM, n ϭ 8);
addition to the superfusing ACSF with that recorded at the peak of the
CD1, 1.87 Ϯ 0.17 spikes/sec (n ϭ 7); C57BL6, 1.49 Ϯ 0.11 spikes/
action of the drug, i.e., 3–10 min after starting the drug infusion. When
sec (n ϭ 12), and 5-HTTϩ/ϩ, 1.89 Ϯ 0.15 spikes/sec (n ϭ 12).
an agonist was applied in the presence of an antagonist, the effect of the
Furthermore, the baseline firing rate of DRN 5-HT neurons was
agonist was compared with the baseline firing rate and with the discharge
frequency recorded during superfusion with the antagonist alone.
also not significantly different from these values in heterozygous
Intracellular recordings of serotoninergic neurons in the dorsal raphe
5-HTTϩ/Ϫ, 1.88 Ϯ 0.19 spikes/sec (n ϭ 6) and homozygous
nucleus and pyramidal neurons in the hippocampus. 5-HT neurons in the
5-HTTϪ/Ϫ mutants, 1.66 Ϯ 0.18 spikes/sec (n ϭ 10). 2180 J. Neurosci., March 15, 2001, 21(6):2178–2185
Receptors in 5-HT Transporter Knock-Out Mice
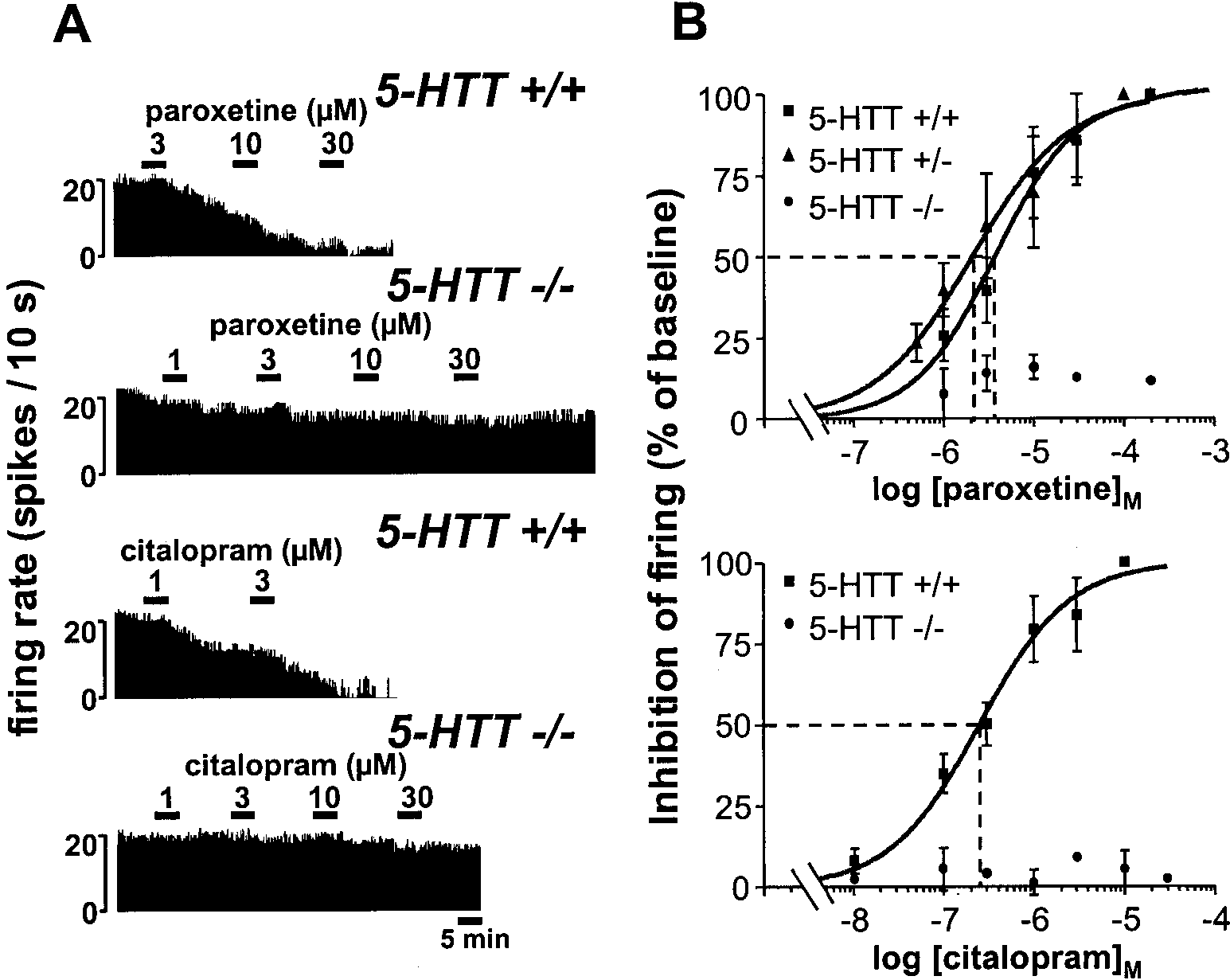
Figure 1. Effects of paroxetine and
and wild-type mice. A, Integrated firing
centrations of paroxetine (top) or cita-
lopram (bottom) on the electrical activ-
mice. B, Concentration-dependent inhi-
bition by paroxetine (top) or citalopram
(bottom) of the firing of DRN 5-HT neu-
induced inhibition is expressed as a per-
centage of the baseline firing rate. Each
cells. The dotted lines illustrate the deter-
Effects of the SSRIs paroxetine and citalopram
Ipsapirone also inhibited the discharge of DRN 5-HT cells in
Like that previously reported under similar conditions in rats (Le
the 5-HTTϩ/Ϫ and 5-HTTϪ/Ϫ mutants, but within higher con-
Poul et al., 1995), addition of increasing concentrations of parox-
centration ranges than in wild-type animals. Although the in-
etine into the ACSF superfusing brainstem slices resulted in a
crease in the EC50 value of ipsapirone in heterozygous
1A receptor-mediated concentration-dependent inhibition
50 ϭ 115.3 Ϯ 7.3 nM; n ϭ 9) was not
of the firing of DRN 5-HT neurons in wild-type 5-HTTϩ/ϩ mice
significant, that in homozygous 5-HTTϪ/Ϫ mutants (EC50 ϭ
(Fig. 1). A similar effect was noted in heterozygous 5-HTTϩ/Ϫ
3.5 Ϯ 1.1 M; n ϭ 10) was highly significant ( p Ͻ 0.001),
indicating an ϳ55-fold decrease in the potency of the 5-HT
2.10 Ϯ 0.43 M (mean Ϯ SEM, n ϭ 7), did not significantly differ
receptor agonist in the latter group compared with wild-type
from that in wild-type controls, 3.70 Ϯ 0.73 M (n ϭ 7) (Fig. 1).
controls. In spite of these differences, complete blockade of the
By contrast, the same treatment applied to brainstem slices from
discharge of DRN 5-HT cells could be achieved in the three
5-HTTϪ/Ϫ mice produced only a minor (less than or equal to
groups, but with different concentrations of ipsapirone (1 M in
Ϫ15%), concentration-independent, reduction in the firing rate
5-HTTϩ/ϩ and 5-HTTϩ/Ϫ mice, 100 M in 5-HTTϪ/Ϫ mice)
of DRN 5-HT cells, even at paroxetine concentration as high as
(Fig. 2). Similar results were found with 5-CT as 5-HT1 agonist.
Thus, 5-CT (1 nM to 10 M) induced a concentration-dependent
Similar findings were obtained with citalopram, which potently
decrease in the firing rate of DRN 5-HT neurons with a signifi-
inhibited, in a concentration-dependent manner, the discharge of
cantly ( p Ͻ 0.001) lower potency in knock-out (EC50 ϭ 52.7 Ϯ 3.6
nM; n ϭ 10) than in wild-type mice (EC
n ϭ 5), but remained essentially inactive in homozygous
10). However, the relative decrease in 5-CT potency in the
5-HTTϪ/Ϫ mutants (less than or equal to Ϫ10% in the firing rate
mutants (by approximately sixfold) was less than that noted for
at 0.1–30 M citalopram) (Fig. 1).
ipsapirone, as illustrated by the shift to the right of concentra-
tion–response curves, which was of much larger amplitude with
Effect of 5-HT1A autoreceptor stimulation
the latter compared with the former agonist (Fig. 2). In any case,
As expected from the stimulation of somatodendritic 5-HT1A
as expected from their mediation through 5-HT1A autoreceptors,
autoreceptors (Haj-Dahmane et al., 1991), the addition of the
the inhibitory effects of both ipsapirone and 5-CT were prevented
5-HT1A receptor agonist ipsapirone into the ACSF superfusing
by the selective 5-HT1A antagonist WAY 100635 (1–3 nM) in both
brainstem slices resulted in a concentration-dependent inhibition
of the firing of DRN 5-HT neurons in wild-type 5-HTTϩ/ϩ mice
(Fig. 2). Similar effects were noted in c129, CD1, and C57BL6
Effects of 5-HT1A receptor blockade by WAY 100635
mice, and the EC50 value of ipsapirone was not significantly
Further characterization of ipsapirone–WAY 100635 interactions
different in these four murine strains: c129, 61.3 Ϯ 6.1 nM
consisted of investigating the concentration-dependent inhibition
(mean Ϯ SEM, n ϭ 9); CD1, 54.1 Ϯ 3.5 nM (n ϭ 9); C57BL6,
of DRN 5-HT neuron firing by ipsapirone in the absence or the
44.9 Ϯ 6.1 nM (n ϭ 9); and 5-HTTϩ/ϩ, 63.1 Ϯ 7.4 nM (n ϭ 10).
presence of a fixed concentration (2 nM) of the 5-HT1A receptor
Receptors in 5-HT Transporter Knock-Out Mice
J. Neurosci., March 15, 2001, 21(6):2178–2185 2181 Figure 2. Concentration-dependent inhibition by ipsapirone or 5-CT of the electrical activity of DRN 5-HT neurons in 5-HTT knock-out and
wild-type mice. Prevention by WAY 100635. A, Integrated firing rate histograms (in spikes per 10 sec) showing the inhibitory effect of ipsapirone
and its prevention by WAY 100635, on the electrical activity of DRN 5-HT cells in 5-HTTϪ/Ϫ and 5-HTTϩ/Ϫ mutants compared with 5-HTTϩ/ϩ
wild-type mice (top). The effect of 5-CT (bottom), and its prevention by WAY 100635, are illustrated in 5-HTTϩ/ϩ and 5-HTTϪ/Ϫ mice.
Histograms are from different neurons. B, Concentration-dependent inhibition by ipsapirone (top) or 5-CT (bottom) of the firing of DRN 5-HT
neurons in 5-HTTϩ/ϩ, 5-HTTϪ/Ϫ, and/or 5-HTTϩ/Ϫ mice. Agonist-induced inhibition is expressed as a percentage of the baseline firing rate.
Each point is the mean Ϯ SEM of data obtained from three to seven individual cells. The dotted lines illustrate the increase in the EC50 values
(abscissa) of ipsapirone and 5-CT in 5-HTTϪ/Ϫ compared with 5-HTTϩ/ϩ mice. *p Ͻ 0.05; **p Ͻ 0.01; ***p Ͻ 0.001 as compared with the
corresponding inhibition in 5-HTTϩ/ϩ and 5-HTTϩ/Ϫ mice.
antagonist. Data in Figure 3 show that WAY 100635 produced a
Intracellular recordings of DRN 5-HT neurons and
shift to the right of the ipsapirone curve in wild-type as well as
hippocampal pyramidal neurons
mutant mice, as expected from competitive inhibition of the effect
of ipsapirone by WAY 100635. Calculation of the IC50 value of
In the absence of drugs, 5-HT cells recorded in 5-HTTϩ/ϩ (n ϭ
WAY 100635 from these curves yielded 0.065 Ϯ 0.015, 0.078 Ϯ
6) as well as in null mutants (n ϭ 5), exhibited similar membrane
0.021, and 0.140 Ϯ 0.035 nM (means Ϯ SEM; n Ն 5 for each value)
potential and Rin ranging from Ϫ66 to Ϫ83 mV and 158 to 255
in 5-HTTϩ/ϩ, 5-HTTϩ/Ϫ, and 5-HTTϪ/Ϫmice, respectively.
M⍀, respectively. Bath-applied ipsapirone evoked both a
In a second series of experiments, the concentration-dependent
concentration-dependent membrane hyperpolarization (with a
prevention by WAY 100635 (5 pM-5 nM) of the inhibitory effect of
maximal response of Ϫ4.50 Ϯ 0.76 mV, n ϭ 4, in the presence of
a fixed concentration of ipsapirone (300 nM) on the discharge of
300 nM ipsapirone) and a decrease in Rin (down to 62% of
DRN 5-HT neurons was compared in 5-HTTϩ/ϩ and
baseline value) of DRN 5-HT neurons in wild-type mice (Fig.
5-HTTϪ/Ϫ mice. Calculations of the IC50 of the 5-HT1A antag-
4A). By contrast, neither the membrane potential nor the Rin of
onist yielded values in the same range as those calculated from
DRN 5-HT neurons in slices from 5-HTTϪ/Ϫ mutants were
the previous series of experiments and did not significantly differ
affected by ipsapirone, even at a concentration as high as 30 M
between wild-type (0.093 Ϯ 0.032 nM; n ϭ 5) and knock-out
(Fig. 4A). Different results were obtained with the other agonist
(0.045 Ϯ 0.014 nM; n ϭ 4) animals.
tested, 5-CT, because this compound, in contrast to ipsapirone,
2182 J. Neurosci., March 15, 2001, 21(6):2178–2185
Receptors in 5-HT Transporter Knock-Out Mice
Figure 3. Competitive inhibition by WAY 100635 of the negative effect of ipsapirone on the firing of DRN 5-HT neurons in 5-HTTϪ/Ϫ and 5-HTTϩ/Ϫ
mutants compared with 5-HTTϩ/ϩ wild-type mice. Experiments were as described in the legend to Figure 2 except that the effects of various
concentrations of ipsapirone were tested in the absence (black symbols) or the presence (open symbols) of 2 nM WAY 100635. Inhibition caused by
ipsapirone is expressed as a percentage of baseline firing rate. Each point is the mean Ϯ SEM of data obtained from five to seven individual cells. *p Ͻ
0.05; **p Ͻ 0.01; ***p Ͻ 0.001 as compared with corresponding data in the absence of WAY 100635.
produced a concentration-dependent membrane hyperpolariza-
well as wild-type (data not shown) mice. Concentration-
tion of DRN 5-HT neurons in both knock-out and wild-type mice
dependent prevention by WAY 100635 (0.3–10 nM) of the mem-
(Fig. 4B,C). As expected from the higher agonist efficacy of 5-CT
brane hyperpolarization induced by 300 nM 5-CT showed no
compared with ipsapirone (Bockaert et al., 1987; Hoyer et al.,
differences between wild-type (IC50 ϭ 0.96 Ϯ 0.04 nM; n ϭ 3) and
1994), maximal membrane hyperpolarization induced by the
knock-out (IC50 ϭ 1.07 Ϯ 0.06 nM; n ϭ 3) mice (Fig. 5C).
former agonist was (ϳ2.5-fold) larger than that observed with the
latter in wild-type animals (Fig. 4C). Concentration-dependent
DISCUSSION
curves showed no difference in 5-CT-induced maximal mem-
The present study showed that the lack of 5-HT reuptake because
brane hyperpolarization in 5-HTTϪ/Ϫ versus 5-HTTϩ/ϩ mice;
of the deletion of exon 2 in the 5-HTT gene (Bengel et al., 1998)
however, the potency of this agonist was significantly ( p Ͻ 0.001)
induces major alterations in central 5-HT neurotransmission. In
50 ϭ 89.6 Ϯ 2.9 nM; n ϭ 3) than in
erts a key role in the modulation of 5-HT tone (Hamon, 1997), is
50 ϭ 11.9 Ϯ 1.0 nM; n ϭ 3) animals (Fig. 4C) In both
groups, the effects of 5-CT (300 nM) were completely prevented
deeply desensitized in the knock-out 5-HTTϪ/Ϫ mice. However,
by WAY 100635 (10 nM), which, on its own, affected neither the
such a functional adaptation does not extend to all 5-HT1A
receptors in brain because those located postsynaptically in the
in of DRN 5-HT cells (Fig. 4 B; data
hippocampus were found to exhibit the same characteristics in
Like that previously reported in rats (Rigdon and Wang, 1991;
Like that noted for DRN 5-HT cells, no significant differences
Le Poul et al., 1995), the increase in extracellular 5-HT concen-
were found in the membrane potential (range, Ϫ53 to Ϫ78 mV)
trations within the DRN of brainstem slices exposed to SSRI
and the Rin value (range, 48–133 M⍀) of CA1 pyramidal cells
(paroxetine, citalopram) was found to trigger a 5-HT1A
between 5-HTTϪ/Ϫ mutant and 5-HTTϩ/ϩ wild-type mice. The
autoreceptor-mediated inhibition of DRN 5-HT cell firing in
addition of increasing concentrations (30 nM to 1 M) of 5-CT
wild-type mice. This response offered a relevant model to further
into the ACSF superfusing hippocampal slices from wild-type
assess the lack of 5-HTT in the knock-out animals, and indeed, as
mice elicited a hyperpolarization of cell membrane (maximal
expected, neither paroxetine nor citalopram were able to produce
response, Ϫ6.55 Ϯ 0.41 mV with 300 nM 5-CT; n ϭ 8) and a
a concentration-dependent inhibition of DRN 5-HT cell firing in
decrease in Rin value (Ϫ44.2% with 300 nM 5-CT) (Fig. 5A,B).
5-HTTϪ/Ϫ mutants. These electrophysiological data further con-
These effects were reversible with recovery of predrug values
firmed previous autoradiographic and biochemical results show-
within ϳ15 min after removal of 5-CT from the superfusing
ing the complete absence of the SSRI molecular target, i.e., the
ACSF (Fig. 5A). As illustrated in Figure 5B, 5-CT-induced effects
5-HTT, in the homozygous mutants (Bengel et al., 1998; Fabre et
were clearly concentration-dependent, with an EC50 value of
al., 2000). In contrast, experiments performed with tissues from
41.0 Ϯ 4.0 nM (n ϭ 6). Similar effects were noted on CA1
heterozygous 5-HTTϩ/Ϫ mice showed that paroxetine inhibited
pyramidal cells in hippocampal slices from 5-HTTϪ/Ϫ animals,
DRN 5-HT cell firing with the same potency in these mutants as
and indeed the maximal hyperpolarization (Ϫ6.05 Ϯ 0.50 mV;
in wild-type animals, although the density of 5-HT transporter
n ϭ 9) and decrease in Rin (Ϫ43.7%) in response to 300 nM 5-CT
binding sites was only half in the former compared with the latter
(Fig. 5), and the potency of this drug (EC50 ϭ 51.1 Ϯ 5.0 nM; n ϭ
group (Fabre et al., 2000). Interestingly, Bengel et al. (1998)
9) to trigger these effects, were not significantly different in the
reported that in vitro synaptosomal [3H]5-HT uptake was also
homozygous mutants compared with wild-type mice.
unchanged in 5-HTTϩ/Ϫ compared with wild-type animals. It
As expected from effects mediated through 5-HT1A receptor
can thus be hypothesized that adaptive changes in 5-HTT intrin-
stimulation, both the membrane hyperpolarization and the de-
sic activity very probably occur to compensate for the (partial)
crease in Rin value caused by 300 nM 5-CT could be completely
loss of 5-HTT protein in heterozygous 5-HTTϩ/Ϫ mutants.
prevented by bath application of 10 nM of the selective 5-HT1A
One of the most interesting observations made in our studies is
receptor antagonist WAY 100635 (Fig. 5A). On its own, WAY
that spontaneous 5-HT neuron firing in brainstem slices was not
100635 affected neither the membrane potential nor the Rin value
altered in 5-HTTϪ/Ϫ mutants compared with wild-type mice.
of CA1 pyramidal neurons in homozygous mutant (Fig. 5A) as
Because the electrophysiological activity of DRN 5-HT cells is
Receptors in 5-HT Transporter Knock-Out Mice
J. Neurosci., March 15, 2001, 21(6):2178–2185 2183 Figure 4. Differential effects of ipsapirone and 5-CT on intracellularly recorded DRN 5-HT neurons in 5-HTT knock-out and wild-type mice. A, Chart
recordings of membrane potential of a DRN 5-HT neuron in a brainstem slice from a 5-HTTϩ/ϩ (top) versus a 5-HTTϪ/Ϫ (bottom) mouse. Each successive
concentration of ipsapirone was applied for 4 min. B, Same as in A, except that 5-CT (300 nM for 4 min) was substituted for ipsapirone. Bottom recording
shows the prevention by 10 nM WAY 100635 of 5-CT-induced hyperpolarization of the same cell as that corresponding to the middle recording. Downward
and upward rapid deflections in A and B are electrotonic cell membrane responses to constant current steps (Ϫ200 to ϩ200 pA) injected through the
recording electrode. Similar data were obtained in at least five cells in each group. C, Concentration–response curves of 5-CT-induced hyperpolarization
of DRN 5-HT neurons in 5-HTTϩ/ϩ and 5-HTTϪ/Ϫ mice. Each point is the mean Ϯ SEM of data obtained in three to five cells for each concentration
of 5-CT. Dotted lines point to the EC50 values (abscissa). **p Ͻ 0.01; *p Ͻ 0.05 compared with respective hyperpolarization in the 5-HTTϩ/ϩ group.
negatively controlled by extracellular 5-HT acting at 5-HT1A
5-HT neuron firing in brainstem slices from mice of various
autoreceptors (Sprouse and Aghajanian, 1987; Haj-Dahmane et
strains, including 5-HTTϩ/ϩ animals. With respect to the inhib-
al., 1991), one would have expected that the lack of 5-HT re-
itory effect of ipsapirone, heterozygous 5-HTTϩ/Ϫ mice did not
uptake produces some reduction in their firing rate because of the
significantly differ from wild-type mice (further supporting the
resulting increase in extracellular 5-HT levels in 5-HTTϪ/Ϫ
idea that compensatory changes occurred in these mutants, see
mice. Indeed, using in vivo microdialysis, marked increases (by at
above), whereas homozygous 5-HTTϪ/Ϫ mutants were much less
least sixfold) in extracellular 5-HT levels were found in the
sensitive to the drug. Indeed ipsapirone potency was ϳ55-fold
substantia nigra (Fabre et al., 2000) and the striatum (Andrews et
lower in the latter animals than in wild-type mice. Similar results
al., 1998) of 5-HTTϪ/Ϫ compared with 5-HTTϩ/ϩ mice. No
were found using 5-CT, except that the potency of this agonist to
data have yet been published concerning extracellular 5-HT con-
inhibit DRN 5-HT neuron firing was decreased by only approxi-
centrations within the DRN, but it can be reasonably assumed
mately sixfold in 5-HTTϪ/Ϫ compared with 5-HTTϩ/ϩ animals.
that they are also markedly enhanced in 5-HTTϪ/Ϫ mice, espe-
This difference between the two agonists was as expected from
cially because the DRN contains a high density of 5-HT reuptake
their respective efficacy at 5-HT1A receptors, because it is well
sites (Bengel et al., 1997; Rattray et al., 1999).
established (Kenakin, 1993) that reductions in receptor number
Because the most probable explanation for the maintenance of
and/or coupling, such as those affecting 5-HT1A autoreceptors in
normal basal firing rate of DRN 5-HT cells in 5-HTTϪ/Ϫ mice is
5-HTTϪ/Ϫ mice (Fabre et al., 2000), decrease to a greater extent
that 5-HT1A autoreceptor-mediated inhibitory control is altered,
the response to a low-efficacy (partial) agonist such as ipsapirone
we directly investigated the functional properties of DRN
(Bockaert et al., 1987), than a high-efficacy (full) agonist such as
5-HT1A autoreceptors in these mutants. Like that previously
observed in rats (Haj-Dahmane et al., 1991), bath application of
Interestingly, in addition to that of 5-HT1A receptor agonists,
ipsapirone induced a concentration-dependent inhibition of DRN
the potency of baclofen, a GABA-B receptor agonist, to inhibit
2184 J. Neurosci., March 15, 2001, 21(6):2178–2185
Receptors in 5-HT Transporter Knock-Out Mice
Figure 5. 5-CT-induced hyperpolariza-
type mice. Prevention by WAY 100635. A, Chart recordings of membrane poten-
min after cessation of 5-CT application.
cording electrode. B, Concentration–re-
concentration of 5-CT. Dotted lines point
to the EC50 values (abscissa). C, Con-
centration–response curves of the antag-
Ϫ5.8 to Ϫ6.7 mV; response range for n ϭ 9 individual cells in each group). Each point is the mean Ϯ SEM of data obtained in three or four cells for
each concentration of WAY 100635. The dotted lines indicate the IC50 values of WAY 100635 (abscissa) against 5-CT-evoked hyperpolarization.
the discharge of DRN 5-HT neurons, was also found to be
rats (Le Poul et al., 1995, 2000). It has to be stressed, however,
decreased in 5-HTTϪ/Ϫ versus 5-HTTϩ/ϩ mice (Mannoury La
that 5-HTT is usually inhibited for only 2–3 weeks in pharmaco-
Cour et al., 2000). Because both 5-HT1A and GABA-B receptors
logical models, whereas it is completely inactivated for the whole
share the same pool of G-proteins (Andrade et al., 1986), it can be
life in knock-out animals, including at critical periods during
inferred that possible alterations in this pool underlie their con-
development when 5-HT can play specific actions on brain mat-
uration (Emerit et al., 1992; Lotto et al., 1999).
Further analyses of the 5-HT1A-mediated responses by intra-
Another key feature of chronic SSRI treatments is the differen-
cellular recording of DRN 5-HT cells could not be performed
tial fate of hippocampal postsynaptic 5-HT1A receptors versus
with ipsapirone because this partial 5-HT1A agonist lost its ca-
DRN 5-HT1A autoreceptors in rats subjected to such treatments.
pacity to hyperpolarize the cell membrane in 5-HTTϪ/Ϫ mu-
Thus, in contrast to the latter receptors, those in the hippocampus
tants. This led us to use the full 5-HT1 agonist, 5-CT, whose
do not desensitize after chronic SSRI administration (Haddjeri et
effects on the membrane potential and Rin also appeared to be
al., 1998; Le Poul et al., 2000). To assess further the possible
completely prevented by WAY 100635, as expected from their
relevance of 5-HTT gene knock-out as a model of chronic 5-HTT
mediation through 5-HT1A receptors (Hamon, 1997). Indeed,
blockade by SSRI, we investigated the functional characteristics of
5-CT was still able to hyperpolarize the plasma membrane of
5-HT1A receptors on pyramidal cells in the CA1 area of the
DRN 5-HT neurons in 5-HTTϪ/Ϫ mice, but with a significantly
hippocampus in 5-HTTϪ/Ϫ mutants compared with wild-type
lower potency than in wild-type animals. The differences between
mice. 5-CT, rather than ipsapirone, was used in these experiments
ipsapirone and 5-CT revealed by these intracellular recording
because the partial agonist properties of the latter ligand produced
experiments were also as expected from respective changes in the
only minor, not reliably measurable, hyperpolarization of CA1
response to a partial and a full agonist (Bockaert et al., 1987;
pyramidal neurons (L. Lanfumey, unpublished observations). Like
Hoyer et al., 1994) after alterations in their shared receptors
that observed in rats (Corradetti et al., 1998), 5-CT application
(Kenakin, 1993) such as those observed in DRN 5-HT1A autore-
onto mouse hippocampal slices produced both a hyperpolarization
of the plasma membrane and a decreased Rin of CA1 pyramidal
Previous studies in rats have shown that chronic impairment of
cells that could be completely prevented by WAY 100635, demon-
5-HT reuptake by long-term SSRI treatment also induces a sig-
strating their mediation through 5-HT1A receptors. Comparison of
nificant desensitization of DRN 5-HT1A autoreceptors (Chaput
the potency of 5-CT to induce these effects in 5-HTTϪ/Ϫ versus
et al., 1986; Jolas et al., 1994; Kreiss and Lucki, 1995; Le Poul et
5-HTTϩ/ϩ mice revealed no difference between the two groups,
al., 1995, 2000), thereby suggesting that similar mechanisms are
indicating that postsynaptic 5-HT1A receptors were not desensi-
responsible for this adaptive phenomenon in both SSRI-treated
tized in the knock-out animals. In line with these observations,
animals and 5-HTT knock-out mice. However, differences also
Fabre et al. (2000) recently reported that 5-HT1A receptor-evoked
exist between these two experimental models because in contrast
[35S]GTP-␥-S specific binding was significantly decreased in the
to that found in 5-HTTϪ/Ϫ mice (Fabre et al., 2000), DRN
DRN but not the hippocampus in 5-HTTϪ/Ϫ compared with
5-HT1A autoreceptors are not downregulated in SSRI-treated
Receptors in 5-HT Transporter Knock-Out Mice
J. Neurosci., March 15, 2001, 21(6):2178–2185 2185
In conclusion, the present in vitro electrophysiological investiga-
distribution of serotoninergic projections from the dorsal raphe nu-
tions demonstrated that somatodendritic 5-HT
Hirst WD, Price GW, Rattray M, Wilkin GP (1998) Serotonin trans-
the DRN, but not postsynaptic 5-HT1A receptors in the hippocam-
porters in adult rat brain astrocytes revealed by [ 3H]5-HT uptake into
pus, are desensitized in knock-out mice that lack 5-HT reuptake
glial plasmalemmal vesicles. Neurochem Int 33:11–22.
capacity. These adaptive changes closely resemble those induced
Hoyer D, Clarke DE, Fozard JR, Hartig PR, Martin GR, Mylecharane
EJ, Saxena PR, Humphrey PPA (1994) VII. International Union of
by chronic SSRI treatment, indicating that 5-HTTϪ/Ϫ mice can be
Pharmacology classification of receptors for 5-hydroxytryptamine (se-
considered as a model to further investigate the molecular mech-
rotonin). Pharmacol Rev 46:157–203.
Jacobs BL, Azmitia EC (1992) Structure and function of the brain sero-
anisms underlying the differential regulation of 5-HT1A autorecep-
tonin system. Physiol Rev 72:165–229.
Jolas T, Haj-Dahmane S, Kidd EJ, Langlois X, Lanfumey L, Fattaccini
CM, Vantalon V, Laporte AM, Adrien J, Gozlan H, Hamon M (1994)
explains why the basal firing rate of DRN 5-HT neurons remains at
ically with a novel antidepressant, cericlamine. J Pharmacol Exp Ther
the same level as in wild-type animals despite marked increases in
extracellular 5-HT levels in the mutants. Whether these differential
Kenakin T (1993) Pharmacologic analysis of drug-receptor interaction,
changes in 5-HT1A receptors in the DRN versus the hippocampus
Kreiss DS, Lucki I (1995) Effects of acute and repeated administration
also account for the behavioral alterations in 5-HTTϪ/Ϫ mice
of antidepressant drugs on extracellular levels of 5-hydroxytryptamine
measured in vivo. J Pharmacol Exp Ther 274:866–876.
(Wichems et al., 2000) should deserve further investigations.
Le Poul E, Laaris N, Doucet E, Laporte AM, Hamon M, Lanfumey L
(1995) Early desensitization of somato-dendritic 5-HT1A autorecep-
tors in rats treated with fluoxetine or paroxetine. Naunyn Schmiede-
REFERENCES
Andrade R, Malenka RC, Nicoll RA (1986) A G protein couples sero-
Le Poul E, Boni C, Hanoun N, Laporte AM, Laaris N, Chauveau J,
tonin and GABAB receptors to the same channels in hippocampus.
Hamon M, Lanfumey L (2000) Differential adaptation of brain
5-HT1A and 5-HT1B receptors and 5-HT transporter in rats treated
Andrews AM, Wichems CH, Li Q, Heils A, Lesch KP, Murphy DL
chronically with fluoxetine. Neuropharmacology 39:110–122.
(1998) A microdialysis study of the effects of high K ϩ and paroxetine
Lesch KP (1997) Molecular biology, pharmacology, and genetics of the
on extracellular serotonin concentrations in serotonin transporter
serotonin transporter: psychobiological and clinical implications. In:
knock-out mice. Soc Neurosci Abstr 24:1112.
Serotoninergic neurons and 5-HT receptors in the CNS. Handbook of
Asberg M, Thoren P, Tra¨ksman L (1976) Serotonin depression in a bio-
experimental pharmacology, Vol 129 (Baumgarten HG, Go¨thert M,
chemical subgroup within the affective disorders. Life Sci 191:478–480.
eds), pp 671–705. Berlin: Springer.
Bengel D, Johren O, Andrews AM, Heils A, Mo¨ssner R, Sanvitto GL,
Li Q, Wichems C, Heils A, Van De Kar LD, Lesch KP, Murphy DL
Saavedra JM, Lesch KP, Murphy DL (1997) Cellular localization and
(1999) Reduction of 5-hydroxytryptamine (5-HT)1A-mediated temper-
expression of the serotonin transporter in mouse brain. Brain Res
ature and neuroendocrine responses and 5-HT1A binding sites in 5-HT
transporter knockout mice. J Pharmacol Exp Ther 291:999–1007.
Bengel D, Murphy DL, Andrews AM, Wichems CH, Feltner D,
Lotto B, Upton L, Price DJ, Gaspar P (1999) Serotonin receptor acti-
Heils A, Mo¨ssner R, Westphal H, Lesch KP (1998) Altered brain
vation enhances neurite outgrowth of thalamic neurons in rodents.
methylenedioxymethamphetamine (“ecstasy”) in serotonin transporter-
Mannoury La Cour C, Froger N, Lesch KP, Hamon M, Lanfumey L
deficient mice. Mol Pharmacol 53:649–655.
(2000) Common transduction mechanisms for 5-HT1A and GABA-B
Bockaert J, Dumuis A, Bouhelal R, Sebben M, Cory RN (1987) Piper-
receptors located on 5-HT neurons: further evidence in 5-HTT knock-
azine derivatives including the putative anxiolytic drugs, buspirone and
out mice. Eur J Neurosci [Suppl 11] 12:20.
ipsapirone, are agonists at 5-HT1A receptors negatively coupled with
Masson J, Sagne´ C, Hamon M, El Mestikawy S (1999) Neurotransmitter
adenylate cyclase in hippocampal neurons. Naunyn Schmiedebergs
transporters in the central nervous system. Pharmacol Rev 51:439–464.
Pickel VM, Chan J (1999) Ultrastructural localization of the serotonin
Chang AS, Chang SM, Starnes DM, Schroeter S, Bauman AL, Blakely
transporter in limbic and motor compartments of the nucleus accum-
RD (1996) Cloning and expression of the mouse serotonin trans-
Rattray M, Michael G, Lee J, Wotherspoon G, Bendotti C, Priestley J
Chaput Y, de Montigny C, Blier P (1986) Effects of a selective 5-HT
(1999) Intraregional variation in expression of serotonin transporter
reuptake blocker, citalopram, on the sensitivity of 5-HT autoreceptors:
messenger RNA by 5-hydroxytryptamine neurons. Neuroscience
electrophysiological studies in the rat brain. Naunyn Schmiedebergs
Rigdon GC, Wang CM (1991) Serotonin uptake blockers inhibit the
Corradetti R, Laaris N, Hanoun N, Laporte AM, Le Poul E, Hamon M,
firing of presumed serotoninergic dorsal raphe neurons in vitro. Drug
Lanfumey L (1998) Antagonist properties of (-)pindolol and WAY
100635 at somatodendritic and postsynaptic 5-HT1A receptors in the rat
Rioux A, Fabre V, Lesch KP, Moessner R, Murphy DL, Lanfumey L,
brain. Br J Pharmacol 123:449–462.
Hamon M, Martres MP (1999) Adaptive changes of serotonin 5-HT2A
Cryan JF, Leonard BE (2000) 5-HT1A and beyond: The role of serotonin
receptors in mice lacking the serotonin transporter. Neurosci Lett
and its receptors in depression and the antidepressant response. Hum
Psychopharmacol Clin Exp 15:113–135.
Sanders-Bush E, Breeding M, Knoth K, Tsutsumi M (1989) Sertraline-
Emerit MB, Riad M, Hamon M (1992) Trophic effects of neurotransmit-
induced desensitization of the serotonin 5-HT2 receptor transmem-
ters during brain maturation. Biol Neonate 62:193–201.
brane signaling system. Psychopharmacology 99:64–69.
Fabre V, Beaufour C, Evrard A, Rioux A, Hanoun N, Lesch KP, Murphy
Sprouse JS, Aghajanian GK (1987) Electrophysiological responses of
DL, Lanfumey L, Hamon M, Martres MP (2000) Altered expression
serotoninergic dorsal raphe neurons to 5-HT1A and 5-HT1B agonists.
and functions of serotonin 5-HT1A and 5-HT1B receptors in knock-out
mice lacking the 5-HT transporter. Eur J Neurosci 12:2299–2310.
Sur C, Betz H, Schloss P (1996) Immunocytochemical detection of the
Graham D, Esnaud H, Habert E, Langer SZ (1989) A common binding
serotonin transporter in rat brain. Neuroscience 73:217–231.
site for tricyclic and nontricyclic 5-hydroxytryptamine uptake inhibitors
Tao-Cheng JH, Zhou FC (1999) Differential polarization of serotonin
at the substrate recognition site of the neuronal sodium-dependent
transporters in axons versus soma-dendrites: an immunogold electron
5-hydroxytryptamine transporter. Biochem Pharmacol 38:3819–3826.
microscopy study. Neuroscience 94:821–830.
Haddjeri N, Blier P, de Montigny C (1998) Long-term antidepressant
Trulson ME, Frederickson CJ (1987) A comparison of the electrophys-
treatments result in a tonic activation of forebrain 5-HT1A receptors.
iological and pharmacological properties of serotonin-containing neu-
rons in the nucleus raphe dorsalis, raphe medianus and raphe pallidus
Haj-Dahmane S, Hamon M, Lanfumey L (1991) K ϩ Channel and
recorded from mouse brain slices in vitro: role of autoreceptors. Brain
5-hydroxytryptamine1A autoreceptor interactions in the rat dorsal
raphe nucleus: an in vitro electrophysiological study. Neuroscience
VanderMaelen CP, Aghajanian GK (1983) Electrophysiological and
pharmacological characterization of serotoninergic dorsal raphe neu-
Hamon M (1997) The main features of central 5-HT1A receptors. In:
rons recorded extracellularly and intracellularly in rat brain slices.
Serotoninergic neurons and 5-HT receptors in the CNS. Handbook of
experimental pharmacology, Vol 129, (Baumgarten HG, Go¨thert M,
Wichems C, Li Q, Andrews A, Lesch KP, Murphy DL (2000) Serotonin
eds), pp 239–268. Berlin: Springer.
transporter knock-out mice show a spontaneous behavioural phenotype
Hensler JG, Ferry RC, Labow DM, Kovachich GB, Frazer A (1994)
of increased “anxiety” and stress responses. Int J Neuropsychopharma-
Quantitative autoradiography of the serotonin transporter to assess the
 2180 J. Neurosci., March 15, 2001, 21(6):2178–2185
2180 J. Neurosci., March 15, 2001, 21(6):2178–2185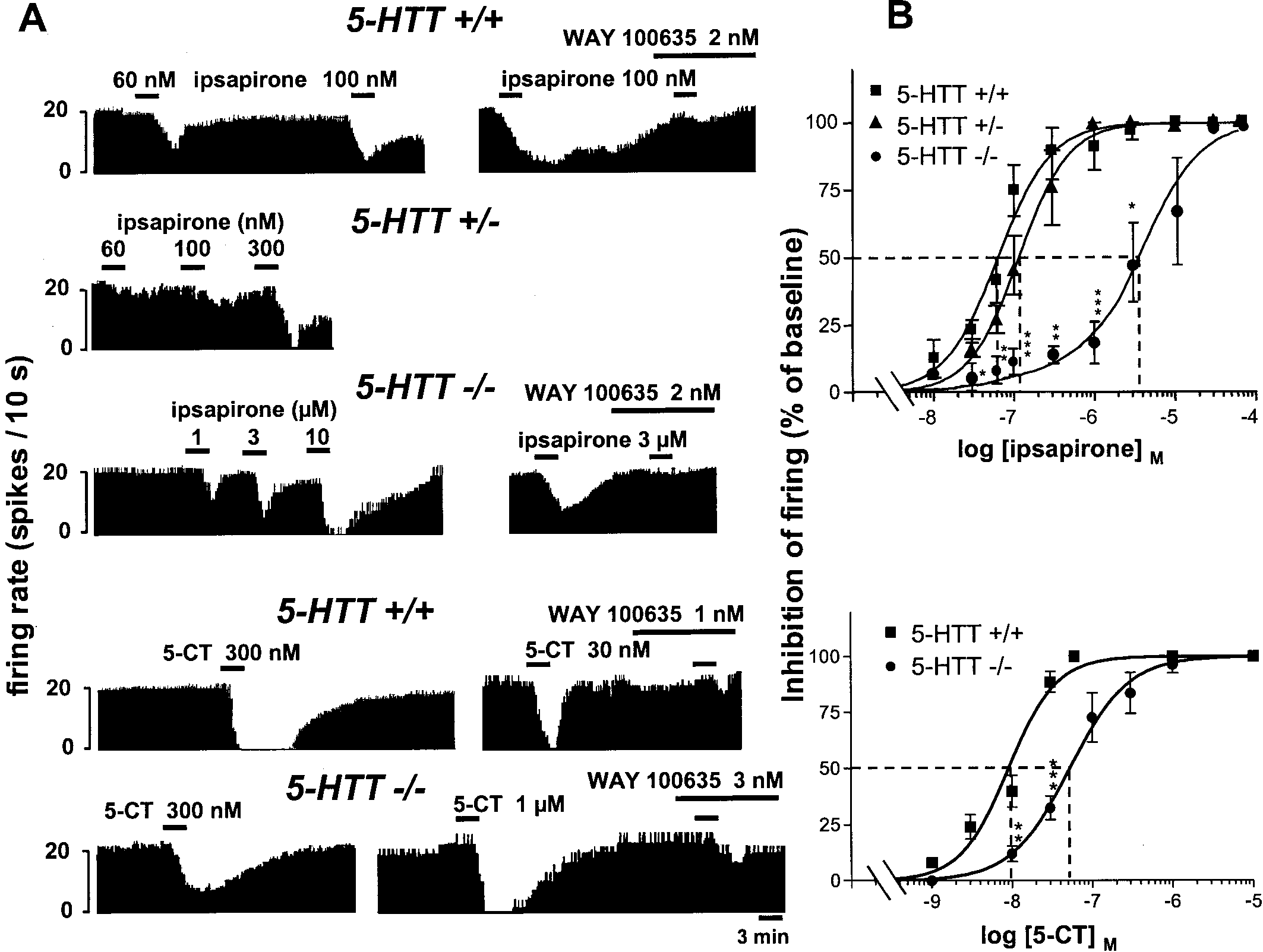 Receptors in 5-HT Transporter Knock-Out Mice
J. Neurosci., March 15, 2001, 21(6):2178–2185 2181
Receptors in 5-HT Transporter Knock-Out Mice
J. Neurosci., March 15, 2001, 21(6):2178–2185 2181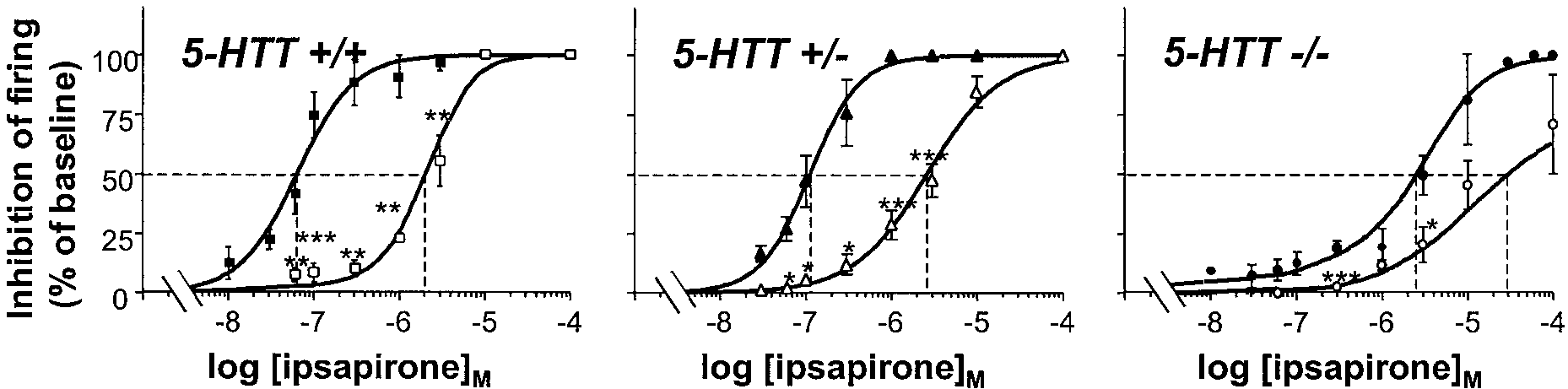 2182 J. Neurosci., March 15, 2001, 21(6):2178–2185
2182 J. Neurosci., March 15, 2001, 21(6):2178–2185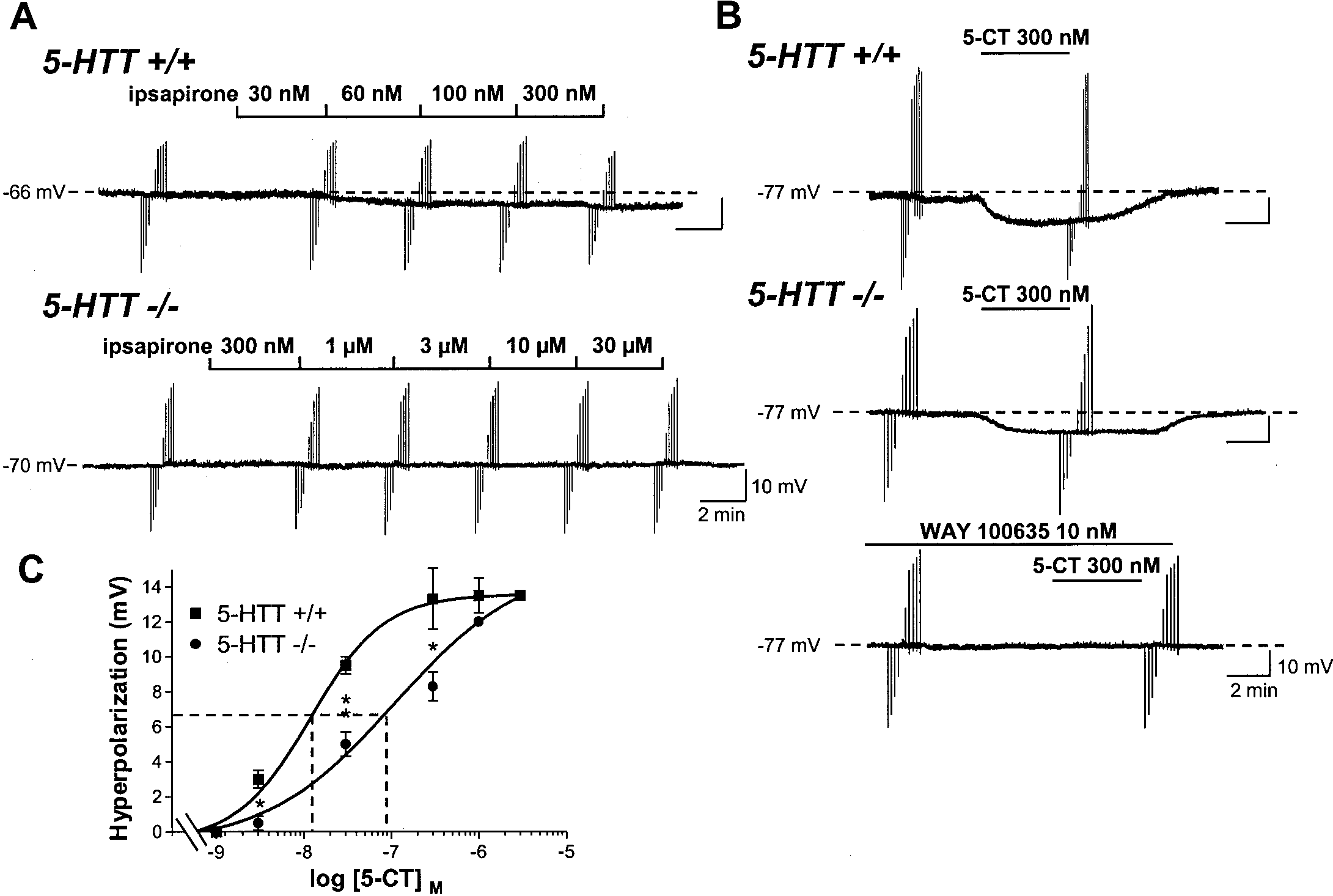 Receptors in 5-HT Transporter Knock-Out Mice
J. Neurosci., March 15, 2001, 21(6):2178–2185 2183
Receptors in 5-HT Transporter Knock-Out Mice
J. Neurosci., March 15, 2001, 21(6):2178–2185 2183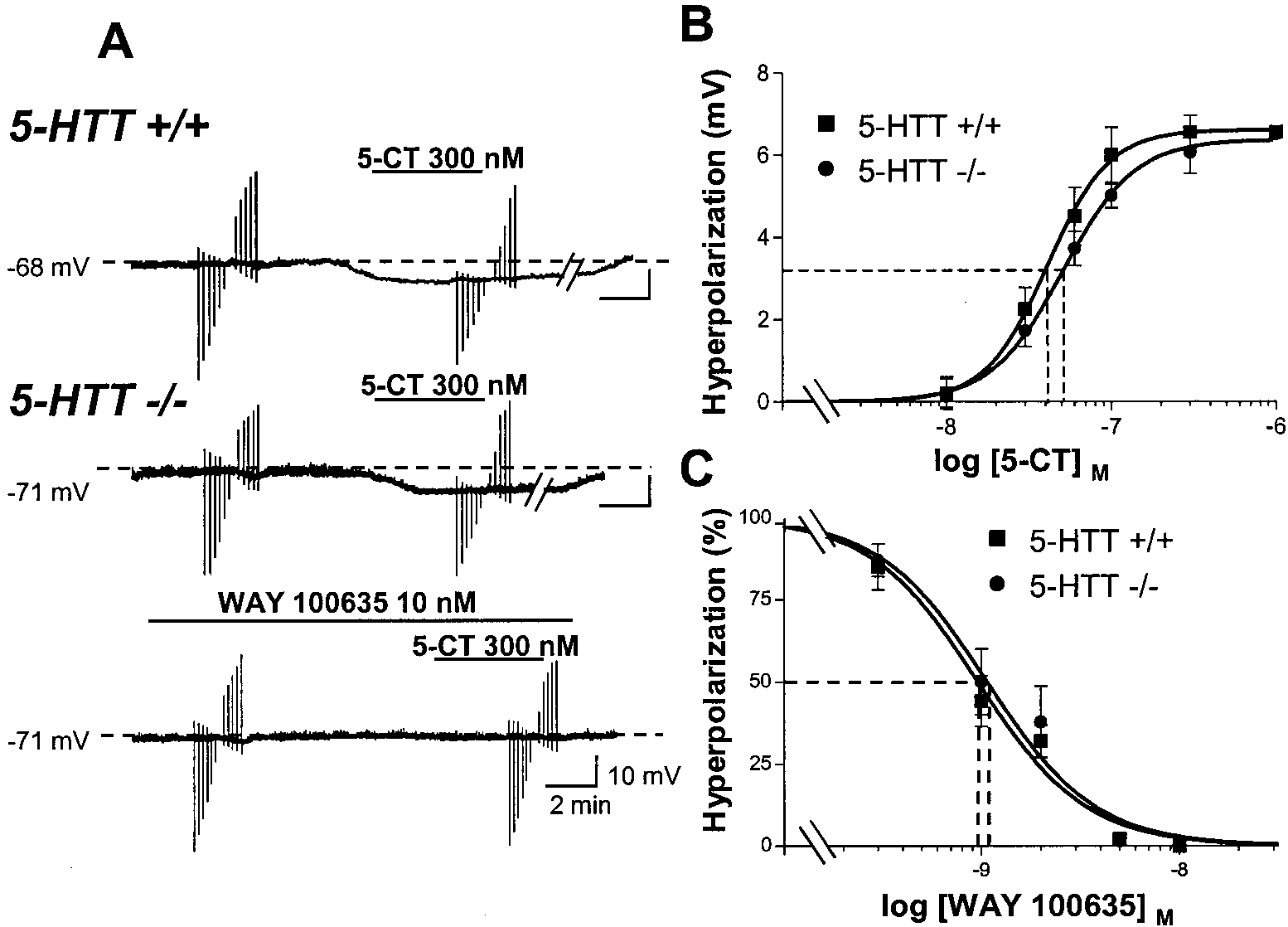 2184 J. Neurosci., March 15, 2001, 21(6):2178–2185
2184 J. Neurosci., March 15, 2001, 21(6):2178–2185