Tadalafil entfaltet seine Wirkung über eine selektive Hemmung der PDE5, wodurch die Konzentration von cGMP im glatten Muskelgewebe stabil bleibt. Diese biochemische Modulation resultiert in einer langanhaltenden Relaxation der Gefäßwände. Der Wirkstoff wird nach oraler Einnahme effizient resorbiert, mit einer Bioverfügbarkeit von rund 80 %. Seine Halbwertszeit von bis zu 36 Stunden ist innerhalb dieser Substanzklasse außergewöhnlich. Abgebaut wird er in der Leber, hauptsächlich durch CYP3A4, mit anschließender biliärer Exkretion. Typische unerwünschte Wirkungen entstehen durch eine verstärkte Vasodilatation, etwa Kopfschmerzen oder Flush. Pharmakologisch wird cialis generika vor allem durch die verlängerte Wirkungsdauer charakterisiert.
Pagesperso.lcp.u-psud.fr
Key role of proline L209 in connecting the distant quinone pockets in the reaction center of Rhodobacter sphaeroides J. Tandori†‡, P. Maroti‡, E. Alexov§, P. Sebban†¶, and L. Baciou†
†Centre de Ge´ne´tique Mole´culaire, Centre National de la Recherche Scientifique, 91198 Gif-sur-Yvette, France; ‡Department of Biophysics, University ofSzeged, H-6722, Szeged, Hungary; and §Howard Hughes Medical Institute and Biochemistry Department, Columbia University, New York, NY 10032
Edited by Hartmut Michel, Max Planck Institute for Biophysics, Frankfurt, Germany, and approved February 25, 2002 (received for review June 28, 2001)
Photosynthetic bacterial reaction centers convert light excitation
side of the complex. The electron is then transferred from QϪA to
into chemical free energy. The initial electron transfer leads to the
a secondary quinone QB within 10–100 s (5–7). Both QA and
consecutive semireductions of the primary (QA) and secondary (QB)
QB are deeply buried within the reaction center protein. The role
quinone acceptors. The Q؊ formations induce proton
of the protein in stabilizing the redox species is essential to
uptake from the bulk. Their magnitudes (H؉͞Q؊ and H؉͞QB ,
ensure high forward electron transfer rates and to prevent charge
respectively) probe the electrostatic interactions within the com-
recombinations to occur. Although chemically identical, QA and
plex. The pH dependence of H؉͞Q؊ A and H؉͞QB were studied in five
QB behave differently. QA, bound to the M subunit in a relatively
single mutants modified at the L209 site (L209P3 F,Y,W,E,T). This
hydrophobic pocket, functions as one electron acceptor and is
residue is situated at the border of a continuous chain of water
never protonated. At variance, QB, bound to the L subunit, is
molecules connecting QB to the bulk. In the wild type (WT), a
surrounded by charged and polar residues and behaves as a
proton uptake band is present at high pH in the H؉͞Q؊ A and H؉͞QB
two-electron gate, accepting sequentially two electrons from QA
curves and is commonly attributed to a cluster of acidic groups
and two protons from the cytoplasm. In chromatophores, the
situated nearby Q B. In the H؉͞QA curves of the L209 variants, this
semiquinone QB can bind a proton below pH 6.8 (8). However,
band is systematically absent but remains in the H؉͞Q؊ B curves.
in isolated RCs, the semiquinones are not directly protonated
Moreover, notable increase of H؉͞Q؊ is observed in the L209
but induce the shift of the pKas of ionizable interacting residues,
mutants at neutral pH as compared with the WT. The large effects
which results in substoichiometric proton uptake by the protein
observed in all L209 mutants are not associated with significant
(9, 10). The proton uptake may occur through a number of water
structural changes (Kuglstatter, A., Ermler, U., Michel, H., Baciou, L.
molecules and protonatable amino acid residues situated be-
& Fritzsch, G. Biochemistry (2001) 40, 4253– 4260). Our data suggest
tween QB and the cytoplasmic surface. Of main interest is
that, in the L209 mutants, the QB cluster does not respond to the
to identify the dynamical and structural role of the protein
A formation as observed in the WT. We propose that, in the
that contributes to the stability of the QA and QB states and
mutants, removal of the rigid proline L209 breaks a necessary
to the energetic and functional connections of their respective
hydrogen bonding connection between the quinone sites. These findings suggest an important role for structural rigidity in ensur-
The partial protonation events that occur on QϪ
ing a functional interaction between quinone binding sites.
formations have been studied by spectroscopic techniques by
using site-directed mutagenesis (9–22) and by numerical meth-
The biological role of bacterial reaction center (RC) mem- ods (23–30). There is a general agreement that the major
brane proteins is to convert light energy into chemical free
response of the protein to the QϪB formation is the change of the
energy. The sequential absorption of two photons by the system
ionization state of acidic residues situated in the QB environ-
results into the production of the doubly reduced and doubly
ment. These residues (L212Glu, L213Asp, L210Asp, and
protonated form of the ultimate electron acceptor of the com-
H173Glu) form a strongly interacting cluster, buffering as a
plex, a ubiquinone (QB). The formed QBH2 molecule then
whole the redox state changes of the quinones. The signature of
delivers its reducing power to the cytochrome bc1 complex,
this cluster is a notable proton uptake band (Ϸ0.8 Hϩ͞QϪB) at low
resulting in the release of protons on the periplasmic side of the
pH and at high pH. The high pH proton uptake band system-
membrane. The resulting transmembrane proton gradient drives
atically disappears in all modified RCs reported so far, where
ATP synthesis through the ATP synthase. The reduction of Q
L212Glu is absent (11, 14, 21), in both the QB and QA states,
coupled to the uptake of protons from the bulk is an important
suggesting that the electrostatic effect of this cluster might be
step shared by many systems involved either in photosynthesis or
extended to the QA environment (12, 13, 15, 18, 31). The long
range electrostatic effect between the QA and QB pocket haS
The three-dimensional structure of the reaction center from
also been proposed on the basis of electrostatic calculations
the purple photosynthetic bacterium Rhodobacter (Rb.) spha-
(24–26, 30, 32, 33). The existence of electrostatic and͞or
eroides is known at atomic resolution (2–4). Three subunits with
conformational-mediated interactions between the two quinone
a total molecular weight of about 100 kDa compose these RCs.
protein pockets of the bacterial reaction centers have been
The transmembrane L and M subunits carry the nine pigments
and cofactors: four bacteriochlorophylls, two bacteriopheophyt-
ins, two ubiquinones 10, and one non-heme iron atom. The third
This paper was submitted directly (Track II) to the PNAS office.
subunit, H, caps the reaction center on the cytoplasmic side of
Abbreviations: P, primary electron donor, a noncovalently linked bacteriochlorophyll
the membrane. The initial photochemical event induced by the
dimer; WT, wild type; L209PY, Pro L209 3 Tyr; L209PF, Pro L209 3 Phe; L209PE, Pro L209 3
absorption of a photon is the creation of the singlet excited state
Glu; L209PW, Pro L209 3 Trp; L209PT, Pro L209 3 Thr; QB and QA, primary and secondary
of a dimer of bacteriochlorophylls (P3P*), which constitutes the
quinone; Rb., Rhodobacter; RC, reaction center; Hϩ͞QA and Hϩ͞QB , proton uptake stoi-chiometries induced by the Q
primary electron donor. P* is a strong reducing species that
¶To whom reprint requests should be addressed. E-mail: sebban@cgm.cnrs-gif.fr.
initiates the electron transfer reaction through the protein. In
about 200 ps, the charge separation occurs between P and the
The publication costs of this article were defrayed in part by page charge payment. Thisarticle must therefore be hereby marked “advertisement” in accordance with 18 U.S.C.
first quinone electron acceptor, QA, situated on the cytoplasmic
§1734 solely to indicate this fact. 6702– 6706 ͉ PNAS ͉ May 14, 2002 ͉ vol. 99 ͉ no. 10
www.pnas.org͞cgi͞doi͞10.1073͞pnas.092327799
pRK404 were previously described (35). The cells were grown in
Erlenmeyer flasks filled to 50% of the total volume with malate
yeast medium supplemented with kanamycin (20 g͞ml) and
tetracycline (2 g͞ml). The cultures were grown in darkness at
30°C on a gyratory shaker (100 rpm). Biochemical Techniques. Cells from Rb. sphaeroides strains (native
or harboring the mutation at L209 site) were disrupted by
sonication in 20 mM Tris (pH 8) buffer in the presence of DNase
and PMSF (1 mM). The intracytoplasmic membranes were
purified as described in ref. 35. The membrane solubilization was
done first by addition of lauryldimethylamine N-oxide (LDAO;
Fluka) to a final concentration of 0.35% in the presence of 100
mM NaCl. The RCs were extracted by a second addition of
LDAO to a final concentration of 0.8% in similar conditions.
The solubilized RCs were subsequently purified on a DEAE
Sepharose CL-6B (Pharmacia) column and eluted at an ionic
strength equivalent to 250 mM NaCl. The ratio of absorbance at
280͞802 nm was in the range 1.5–1.8 for all RC preparations. Proton Uptake Measurements. The RCs were extensively dialyzed
against 50 mM NaCl, 0.03% Triton X-100 during 36 h at 4°C.
Under these conditions, the Tris buffer concentration was kept
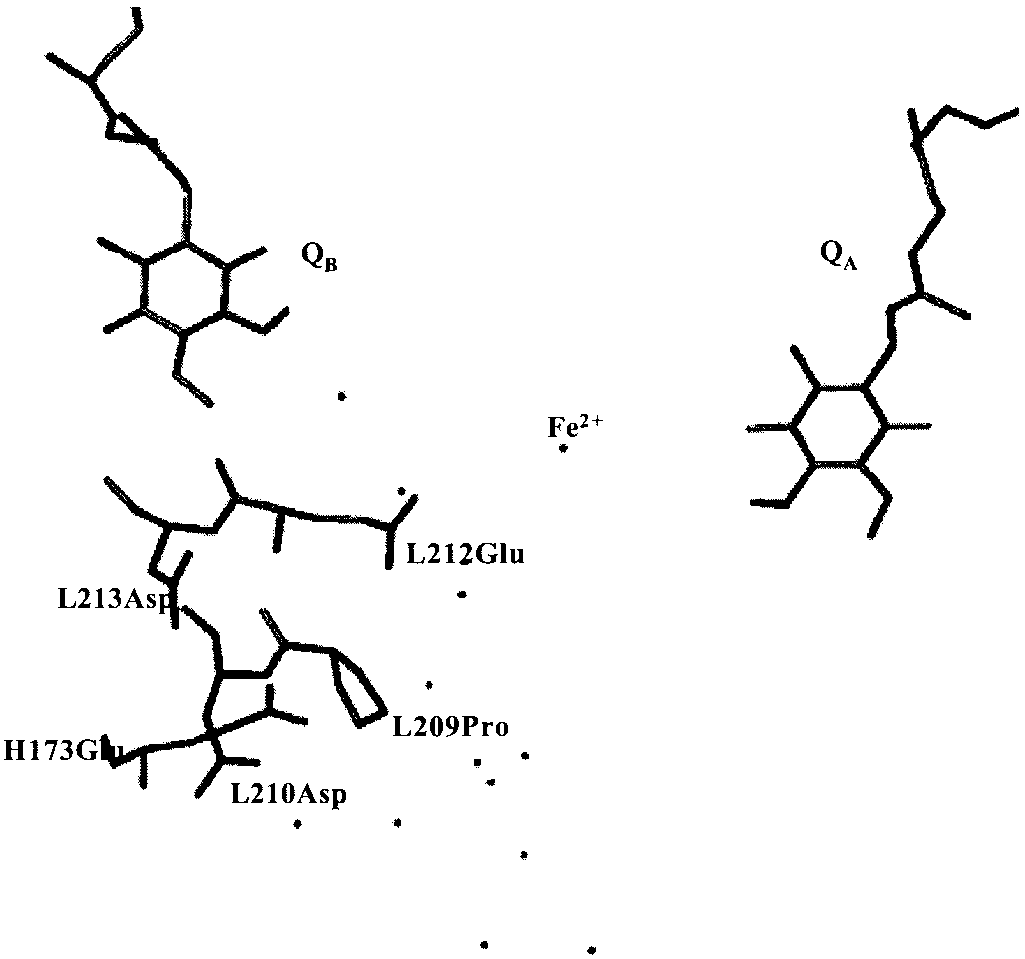
Rb. sphaeroides RC structure showing the two quinones Q
below 10 M. The proton uptake by the RCs was measured on
the QB cluster of acidic residues, and the L209P mutation site. Water molecules
a home made spectrophotometer by following the absorption
connecting QB to the bulk are also represented. Coordinates were taken from
changes at 585 nm of pH sensitive dyes after one saturating (Yag)
laser flash at 532 nm (14). The final proton uptake signal was
obtained with subtracting the buffered sample from the unbuf-
fered signal. The proton uptake by the RCs (Ϸ2 M) was
evoked in experimental works (34–41). The three-dimensional
measured at room temperature in the presence of 20 M
structure of the protein reveals a large hydrogen bond network
bromocresol purple, o-cresol red, or o-cresol-phthaleine, de-
in the quinone proteic region involving numerous ionizable,
polar residues and water molecules (2, 3, 42, 43). It is therefore
The proton uptake stoichiometries were measured in the
of particular interest to investigate to what extent this widely
spread out hydrogen bond network is involved in balancing the
M ferrocene as electron donor to Pϩ and 300
BIOPHYSICS
proton concentration over the key amino acid residues in the two
M ferrocyanide. The calibrations were performed by additions
of known amounts of HCl (1 M stock; Merck). The proton
We report here proton uptake measurements on QϪ
uptake stoichiometries because of the formation of QA were
formations in RC mutants from Rb. sphaeroides in which
measured in the presence of terbutryn (100 M), which prevents
L209Pro has been changed by site-directed mutagenesis to
threonine (L209PT), tryptophane (L209PW), glutamate
The proton uptake by the PQAQB state (⌬HQAQBϪ) is deduced
(L209PE), phenylalanine (L209PF), and tyrosine (L209PY).
from the measured value after one flash (⌬Hobs) according to the
L209Pro is situated at the border of a chain of hydrogen-bonded
water molecules (Fig. 1) that connects QB to the cytoplasmic
⌬H ϩ Ϫ ͓␦ ϩ ␣͑1 Ϫ ␦͔͒⌬H ϩ
surface of the RC (2, 3). Our previous reports concerned the
characterization of the functional properties of these mutants
(35, 44). The x-ray structure of three of these variant proteins
(L209PF, L209PY, and L209PE) has also been determined (45).
⌬HϩϩQAϪ is the proton uptake by the RC in the absence of QB.
The amino acid exchange in the L209PE and L209PT mutants
The fraction of RCs without QB activity (␦) and the partition
functionally mimics the kinetics of the wild-type (WT) RCs (44).
In the L209PE reaction center, the structure remains unchanged
determined from the PϩQϪB 3 PQB charge recombination
compared with the WT structure, except the introduced carbox-
kinetics monitored at 430 nm. Depending on the strain, ␣ varied
ylic side chain of GluL209 located within the water chain (45).
from 0.02 to 0.5 and ␦ from 0.03 to 0.5 as pH is increased from
In the L209PW, L209PY, and L209PF variants, the spectroscopic
6 to 10. The occupancy of the QB site was routinely restored by
analysis suggested a modification of the network of hydrogen
the addition of 60 M ubiquinone-6 (UQ6).
bonds (35, 44). Consistently, the three-dimensional structures of
the L209PF and L209PY mutant RCs show that the mutations
have induced local structural changes of three amino acid
The Stoichiometries of Proton Uptake in the Q؊ A State (H؉͞QA ). The
residues (AspL213, ThrL226, and GluH173) and more distantly
pH titrations of the Hϩ͞QϪA curves measured in the L209PT,
have affected the QB position (45). Despite these different
L209PW, L209PE, L209PY, and L209PF mutants in the pH
structural changes, the similar proton uptake patterns measured
range 6–10.2 are shown in Fig. 2, together with the WT data. The
here for all mutants suggest a crucial role for proline L209 in
WT Hϩ͞QϪA stoichiometry displays a notable proton uptake
connecting both quinone environments.
(Ϸ0.30 Hϩ͞QϪA) at neutral pH and a significant proton uptake
band (Ϸ0.45 Hϩ͞QϪA) at high pH, centered at pH 9. In the L209
Materials and Methods
mutants, below pH 8, the Hϩ͞QϪA proton uptake curves are
Bacterial Strains and Growth Conditions. The design of the Rb.
superimposable to that of the WT, within the experimental
sphaeroides WT or mutant strains harboring pufL mutation on
error. However, above pH 8, any of the introduced mutated side
PNAS ͉ May 14, 2002 ͉ vol. 99 ͉ no. 10 ͉ 6703
pH dependence of the stoichiometries of proton uptake by the PQϪ
pH dependence of the stoichiometries of proton uptake by the PQϪ
state in RCs of the WT (■), the L209PE (ᮀ), the L209PT (ƒ), the L209PY(E), the
state in RCs of the WT (■), the L209PE (ᮀ), the L209PT (ƒ), the L209PY(E), the
L209PW (छ), and the L209PF (‚) mutants. Conditions: Ϸ2 M RCs, 0.03%
L209PW (छ), and the L209PF (‚) mutants. Same conditions as in Fig. 2, except:
Triton X-100, 100 M ferrocene, 300 M ferrocyanide, 50 mM NaCl, 100 M
60 M ubiquinone-6 (UQ6) and no terbutryn present. The error bars reflect the
terbutryn, 20 M dye (bromocresol purple, o-cresol red, or o-cresol-
respective experimental error of each set of measurements.
phthaleine, depending on the pH). The error bars reflect the respectiveexperimental error of each set of measurements.
reduction of either of the two quinone electron acceptors (QA
chains at position L209 cancels the high pH band observed in the
and QB) is an intrinsic observable of the electrostatic interactions
associated with the redox function of the RC.
The high pH proton uptake band is commonly observed in the
WT RCs from Rb. sphaeroides and Rb. capsulatus either on the
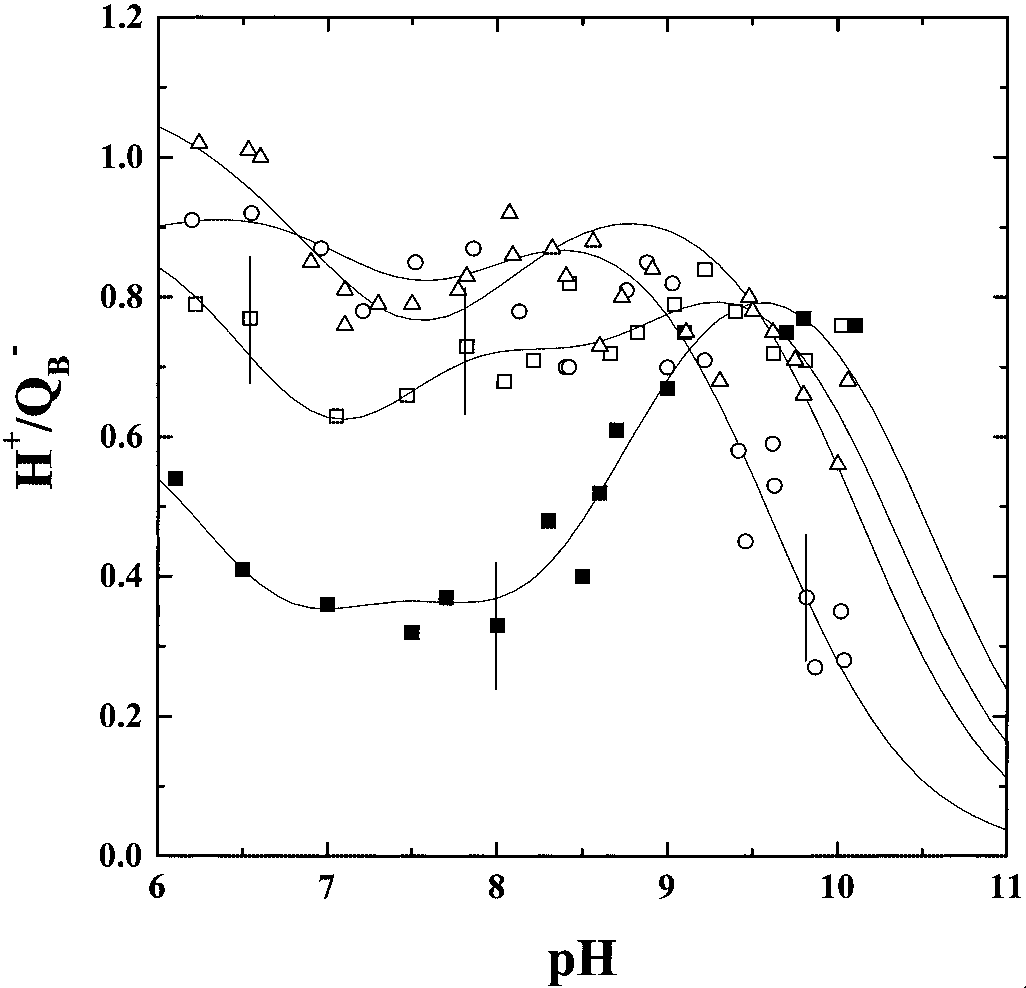
The Stoichiometries of Proton Uptake in the Q؊ B State (H؉͞QB ). Fig.
3 shows the pH titration curves of the Hϩ͞QϪ
QA or QB formation. This band disappears concomitantly in the
the WT and in the L209PE, L209PF, and L209PY mutants. The
Hϩ͞QA and Hϩ͞QB curves in all mutants reported so far in
which L212Glu (situated in the QB pocket at more than 15Å from
B measurements are focused on the RCs from these three
mutants because their three-dimensional structures are available
QA) was changed to a non-protonatable residue (11–14, 21). This
high pH proton uptake band has been attributed to a change in
B stoichiometry is notable (Ϸ0.35– 0.55
the ionization state of L212Glu (11, 14, 18, 21). However, it is
B ) at neutral pH. A significant proton uptake band (Ϸ0.80
likely that this band more generally results from the cluster of
Hϩ͞QϪB) is observed at high pH, centered around pH 9.7.
strongly interacting acidic groups (L212Glu, L213Asp, L210Asp,
In the L209PE mutant, above pH 9, the Hϩ͞QϪB proton uptake
curve is superimposable to that of the WT. However, at lower
networks (12). Consistently, the AspL2133Asn substitution
pH, the Hϩ͞QϪB value is significantly higher than in the WT.
displaces also this band to lower pH (11). The presence of a high
Indeed, a value of about 0.60–0.70 Hϩ is measured down to pH
pH band is due to the cumulative effects of the strong pair-wise
7, below which the proton uptake increases to about 0.80 Hϩ at
interactions within the cluster. Removing any member of the
cluster (as observed when one acidic residue is changed to a
In the L209PF mutant, above pH 9, the Hϩ͞QϪB value is very
non-ionizable residue) shifts to lower pH the highest pKa of both
similar, within the experimental error, to that measured in the
Hϩ͞QϪ and Hϩ͞QϪ curves, resulting into the apparent disap-
WT or in the L209PE mutant. At neutral pH, the Hϩ͞QϪ
pearance of the high pH signature of the cluster (P.S., L.B., and
stoichiometry is higher than in the L209PE mutant: 0.80 Hϩ͞QϪB
are taken up in the pH range 7–9. This value increases up to 1.00
We show here that all L209 mutations specifically suppressed
the high pH proton uptake band on Q (Fig. 2) formation but
In the L209PY mutant, a similar pattern to the L209PF mutant
not on QϪ formation (Fig. 3). Therefore, a complete under-
is measured in the pH range 6–9, with a significant high proton
standing of the observed effects on both Hϩ͞QϪ
uptake (Hϩ͞QϪB Ϸ 0.80–0.90). However, above pH 9, the proton
uptake stoichiometries requires a more complex representation.
uptake values drop, Hϩ͞QϪB being equal to Ϸ0.80 at pH 9, but
Protonation Events Triggered by the Q؊ A Formation. The high pH
band in the Hϩ͞QϪA stoichiometries is absent in the five mutants
Discussion
lacking L209Pro. The crystal structures of the L209PE, L209PY,
In the present paper, we have measured the proton uptake
and L209PF variants have been determined (45). In the crys-
stoichiometries of RC mutants from Rb. sphaeroides in which
tallographic structure of the three variants, no changes in the
L209Pro has been changed by site-directed mutagenesis to
protein backbone were observed, compared with the WT RC
threonine, tryptophane, glutamate, phenylalanine, and tyrosine.
structure. The structural models of the variants show some
The magnitude of proton uptake induced by the one-electron
structural modifications specific to each point-mutation. The
6704 ͉ www.pnas.org͞cgi͞doi͞10.1073͞pnas.092327799
Scheme representing the response of the protein to the formation of QϪ
A and QB in the WT and in the L209P mutants. In the WT, the QA state triggers
the uptake of substoichiometric protons by the cluster from the outside of the protein. In the case of the L209 mutants, the absence of L209Pro softens the protein,altering the connecting relays between the QϪ
A and the QB environment. In the QB state, in the mutants, a substantial additional amount of protons is taken from
the bulk as a consequence of the disorganization of the hydrogen bond network.
structure of the L209PE mutant RC is superimposable to that of
the WT except for the introduced glutamate side chain, which
regard to the high pH band suggests that, in the L209 mutants,
BIOPHYSICS
points toward the hydrogen-bonded water molecules (45) that
B cluster to respond to the QA formation is
connect QB to the cytoplasmic surface of the RC (2, 3). In the
altered (Fig. 4). The rigid side chain of the proline might be of
structure of the L209PY and L209PF mutants, both aromatic
importance for the conformational coupling between the two
side chains are oriented away from the water chain and displace
quinone environments. The absence of L209Pro may soften this
three surrounding side chains (L213Asp, L226Thr, and
coupling, damp the conformational changes, and prevent its
H173Glu) by up to 2.6 Å (45). In the structure of the L209PY
propagation into the QB site. Therefore, the protein dynamics
variant, QB is shifted by Ϸ4 Å and is now located at a position
appear to be critical to ensure the connection between the Q
similar to that reported for the WT reaction center under
illumination (2, 3). In the L209PF variant, the electron density
In the L209PY, -F, and -E mutants, the major effect observed
map reveals an intermediate QB position between the binding
in the QϪ state is an increased proton uptake (0.6–1.0 Hϩ͞QϪ),
sites of the WT protein in the dark and that of the L209PY
as compared with the WT (0.4 Hϩ͞QϪ) below pH 9. The highest
protein. In the L209PE reaction center, the binding site of Q
effect is observed in the L209PY and L209PF mutants.
remains unchanged compared with the WT structure (2, 3).
A proposed mechanistic model to explain the amplitude of
These different structural effects, but resulting into similar pH
proton uptake in the RCs takes into account the movement of
lead us to conclude that the absence of ProL209 per se—and not
QB from the distal position in its neutral state to the proximal
the introduced specific side chain—is responsible for the absence
position in the QB state (46, 47). According to this hypothesis,
of the high pH band observed in the Hϩ͞QϪ
QB in the proximal position is bound via hydrogen bond to
GluL212, and stabilizes thereby the protonated form of the
Protonation Events Triggered by the Q؊
B in the distal position is likely to favor the
B Formation. In the L209PY,
-F, and -E mutants, we do not observe the concomitant drop of
anionic form of GluL212 as suggested by the calculations (28).
Then the amount of protons that is bound at neutral and alkaline
pH is expected to correspond to the fraction of Q
the L209PY mutant, we observe a slight acidic shift of the
position. However, in the L209PY mutant, where QB is found in
the proximal position in its neutral state (45), the same high
B high pH band (Fig. 3), as compared with the WT. This
shift may be correlated to the observed position of Q
amount of proton is taken up as in the L209PF and L209PE
structure of this RC variant in its neutral state, which is in the
variants for which QB is observed in an intermediate or in a
proximal position to the non-heme iron (45). This position has
WT-like position, respectively (45). This result does not support
been suggested to require GluL212 and AspL213 to be proton-
the above proposed mechanistic model. However, in the
ated (28). This result would in turn reduce the strength of the
L209Tyr mutant, the QB ‘‘proximal’’ position may be tilted by
interactions within the cluster in the L209PY mutant, consis-
180° compared with its position in the WT. It could then be that
tently with the observed acidic shift of the Hϩ͞QϪB curve.
the terms ‘‘proximal’’ and ‘‘distal’’ should not mean (as it is in the
PNAS ͉ May 14, 2002 ͉ vol. 99 ͉ no. 10 ͉ 6705
WT) the presence and absence of H-bond between QB and
part) by conformational coupling. The current resolutions of the
GluL212, respectively, in the special cases of the L209 mutations.
three-dimensional structures of the L209 mutants do not provide
It has previously been suggested that, at neutral pH, i.e., in the
any structural changes, explaining for the likely modified dy-
region where we observe a notable increase of the Hϩ͞QϪB values
namics of the protein. Fourier transform infrared spectroscopy
in the mutants, protein surface groups are responsible for the
that may investigate the global vibration modes of these net-
proton binding at the first flash (48, 49). That result would not
support the involvement of the ionization state of GluL212 in the
works, as well as molecular dynamics calculations, will help to
observed phenomenon. In fact, the proton uptake is determined
identify the modified interactions in the mutants.
on a time average base of the exposure of the groups to the
aqueous solution (40). If this mobility is favored by removal of
We thank Marilyn Gunner and Jeroˆme Lavergne for stimulating dis-
the proline from the structure, then the exposure on a time-
cussions, and Tania Bizouarn for careful reading of the manuscript. E.A.
thanks Barry Honig for the support during this work. This work was
supported by the Centre National de la Recherche Scientifique. J.T. was
Conclusion
in part supported by a BALATON grant [Hungarian͞French Ministe`re
The present paper provides evidence that interactions between
des Affaires Etrange`res (No. 00834)], and by a North Atlantic Treaty
the QϪA state and the environment of QB is mediated (at least in
Organization collaborative research grant (LST.CLG 975754).
1. Cramer, W. A. & Knaff, D. B. (1990) in Energy Transduction in Biological
25. Alexov, E., Miksovska, J., Baciou, L., Schiffer, M., Hanson, D. K., Sebban, P. Membranes: A Textbook of Bioenergetics, ed. Cantor, C. R. (Springer, New
& Gunner, M. R. (2000) Biochemistry 39, 5940–5952.
26. Alexov, E. G. & Gunner, M. R. (1999) Biochemistry 38, 8253–8270.
2. Ermler, U., Fritzsch, G., Buchanan, S. K. & Michel, H. (1994) Structure 2,
27. Beroza, P., Fredkin, D. R., Okamura, M. Y. & Feher, G. (1995) Biophys. J. 68,
3. Stowell, M. H., McPhillips, T. M., Rees, D. C., Soltis, S. M., Abresch, E. &
28. Grafton, A. K. & Wheeler, R. A. (1999) J. Phys. Chem. B 103, 5380–5387.
Feher, G. (1997) Science 276, 812–816.
29. Rabenstein, B., Ullmann, G. M. & Knapp, E. W. (1998) Biochemistry 37,
4. McAuley, K. E., Fyfe, P. K., Ridge, J. P., Cogdell, R. J., Isaacs, N. W. & Jones,
M. R. (2000) Biochemistry 39, 15032–15043.
30. Lancaster, C. R., Michel, H., Honig, B. & Gunner, M. R. (1996) Biophys. J. 70,
5. Li, H. & Sherman, L. A. (2000) J. Bacteriol. 182, 4268–4277.
6. Li, J., Gilroy, D., Tiede, D. M. & Gunner, M. R. (1998) Biochemistry 37,
31. Miksovska, J., Maro´ti, P., Schiffer, M., Hanson, D. K. & Sebban, P. (1995) in
Photosynthesis: From Light to Biosphere, ed. Mathis, P. (Kluwer, Dordrecht, The
7. Tiede, D. M., Vazquez, J., Cordova, J. & Marone, P. A. (1996) Biochemistry 35,
32. Brzezinski, P., Okamura, M. Y. & Feher, G. (1992) in The Photosynthetic
8. Lavergne, J., Matthews, C. & Ginet, N. (1999) Biochemistry 38, 4542–4552. Bacterial Reaction Center II, ed. Vermeglio, J. B. A. (Plenum, New York), pp.
9. Maroti, P. & Wraight, C. A. (1988) Biochim. Biophys. Acta 934, 329–347.
10. McPherson, P. H., Okamura, M. Y. & Feher, G. (1988) Biochim. Biophys. Acta
33. Tiede, D. M. & Hanson, D. K. (1992) Protein Relaxation Following Quinone934, 348–368. Reduction in Rhodobacter Capsulatus: Detection of Likely Protonation-Linked
11. Brzezinski, P., Paddock, M. L., Okamura, M. Y. & Feher, G. (1997) Biochim.Optical Absorbance Changes of the Chromatophores (Plenum, New York). Biophys. Acta 1321, 149–156.
34. Ginet, N. & Lavergne, J. (2001) Biochemistry 40, 1812–1823.
12. Tandori, J., Baciou, L., Alexov, E., Maroti, P., Schiffer, M., Hanson, D. K. &
35. Baciou, L. & Michel, H. (1995) Biochemistry 34, 7967–7972.
Sebban, P. (2001) J. Biol. Chem. 276, 45513–45515.
36. Wraight, C. A. & Stein, R. R. (1980) FEBS Lett. 113, 73–77.
13. Miksovska, J., Maro´ti, P., Tandori, J., Schiffer, M., Hanson, D. K. & Sebban,
37. Wraight, C. A. (1979) Biochim. Biophys. Acta 548, 309–327. 35, 15411–15417.
14. Miksovska, J., Ka´lma´n, L., Schiffer, M., Maro´ti, P., Sebban, P. & Hanson, D. K.
38. Prince, R. C. & Dutton, P. L. (1978) in The Photosynthetic Bacteria, eds.
(1997) Biochemistry 36, 12216–12226.
Clayton, R. K. & Sistrom, W. R. (Plenum, New York), pp. 439–453.
15. Miksovska, J., Schiffer, M., Hanson, D. K. & Sebban, P. (1999) Proc. Natl.
39. Prince, R. C. & Dutton, P. L. (1976) Arch. Biochem. Biophys. 172, 329–334. Acad. Sci USA 96, 14348–14353.
40. Maroti, P. & Wraight, C. A. (1997) Biophys. J. 73, 367–381.
16. Nabedryk, E., Breton, J., Okamura, M. Y. & Paddock M. L. (1998) Photosynth.
41. Dutton, P. L., Leigh, J. S. & Wraight, C. A. (1973) FEBS Lett. 36, 169–173. Res. 55, 293–299.
42. Abresch, E. C., Paddock, M. L., Stowell, M. H. B., McPhillips, T. M., Axelrod,
17. Paddock, M. L., Feher, G. & Okamura, M. Y. (1997) Biochemistry 36,
S. M., Soltis, S. M., Rees, D. C., Okamura, M. Y. & Feher, G. (1998) Photosynth.Res. 55, 119–125.
18. Maro´ti, P., Hanson, D. K., Schiffer, M. & Sebban, P. (1995) Nat. Struct. Biol.
43. Lancaster, C. R. & Michel, H. (1997) Structure 5, 1339–1359. 2, 1057–1059.
44. Tandori, J., Sebban, P., Michel, H. & Baciou, L. (1999) Biochemistry 40,
19. Hienerwadel, R., Grzybek, S., Fogel, C., Kreutz, W., Okamura, M. Y., Paddock,
M. L., Breton, J., Nabedryk, E. & Mantele, W. (1995) Biochemistry 34,
45. Kuglstatter, A., Ermler, U., Michel, H., Baciou, L. & Fritzsch, G. (2001)
Biochemistry 40, 4253–4260.
20. Nabedryk, E., Breton, J., Hienerwadel, R., Fogel, C., Ma¨ntele, W., Paddock,
46. Cherepanov, D. A., Bibikov, S. I., Bibikova, M. V., Bloch, D. A., Drachev, L. A.,
M. L. & Okamura, M. Y. (1995) Biochemistry 34, 14722–14732.
Gopta, O. A., Oesterhelt, D., Semenov, A. Y. & Mulkidjanian, A. Y. (2000)
21. McPherson, P. H., Scho¨nfeld, M., Paddock, M. L., Okamura, M. Y. & Feher,
Biochim. Biophys. Acta 1459, 10–34.
G. (1994) Biochemistry 33, 1181–1193.
47. Mulkidjanian, A. Y. (1999) FEBS Lett. 463, 199–204.
22. Takahashi, E. & Wraight, C. A. (1992) Biochemistry 31, 855–866.
48. Turzo´, K., Laczko´, G. & Maro´ti, P. (1998) Photosynth. Res. 55, 235–240.
23. Zachariae, U. & Lancaster, C. R. (2001) Biochim. Biophys. Acta 1505, 280–290.
49. Shinkarev, V. P., Drachev, L. A., Mamedov, M. D., Mulkidjanian, A. J.,
24. Rabenstein, B., Ullmann, G. M. & Knapp, E. W. (2000) Biochemistry 39,
Semenov, A. Y. & Verkhovsky, M. I. (1993) Biochim. Biophys. Acta 1144, 6706 ͉ www.pnas.org͞cgi͞doi͞10.1073͞pnas.092327799
Lorenz il fondatore dell’etologia. La discesa di Maffei Konrad Lorenz toccò l’apice con l’assegnazione delle accuse che provenivano, soprattutto, daicritici francesi rivolte alla cultura italiana, icui rappresentanti, a loro avviso, erano inca-lare sul fenomeno dell’"imprinting"decadenza del teatro appariva alquanto pa-lese, dato che, alle nostre scene, mancava
Embajada de la República Bolivariana de Venezuela en los Emiratos Árabes Unidos Artículo 51: prohíbe a los funcionarios consulares y demás empleados de la oficina ".redactar documento alguno por encargo de los particulares, ni deberán mezclarse en ninguna forma en los contratos y actos de las partes." Los registradores y empleados de su dependencia no podrán solici
 pRK404 were previously described (35). The cells were grown in
Erlenmeyer flasks filled to 50% of the total volume with malate
yeast medium supplemented with kanamycin (20 g͞ml) and
tetracycline (2 g͞ml). The cultures were grown in darkness at
30°C on a gyratory shaker (100 rpm).
pRK404 were previously described (35). The cells were grown in
Erlenmeyer flasks filled to 50% of the total volume with malate
yeast medium supplemented with kanamycin (20 g͞ml) and
tetracycline (2 g͞ml). The cultures were grown in darkness at
30°C on a gyratory shaker (100 rpm).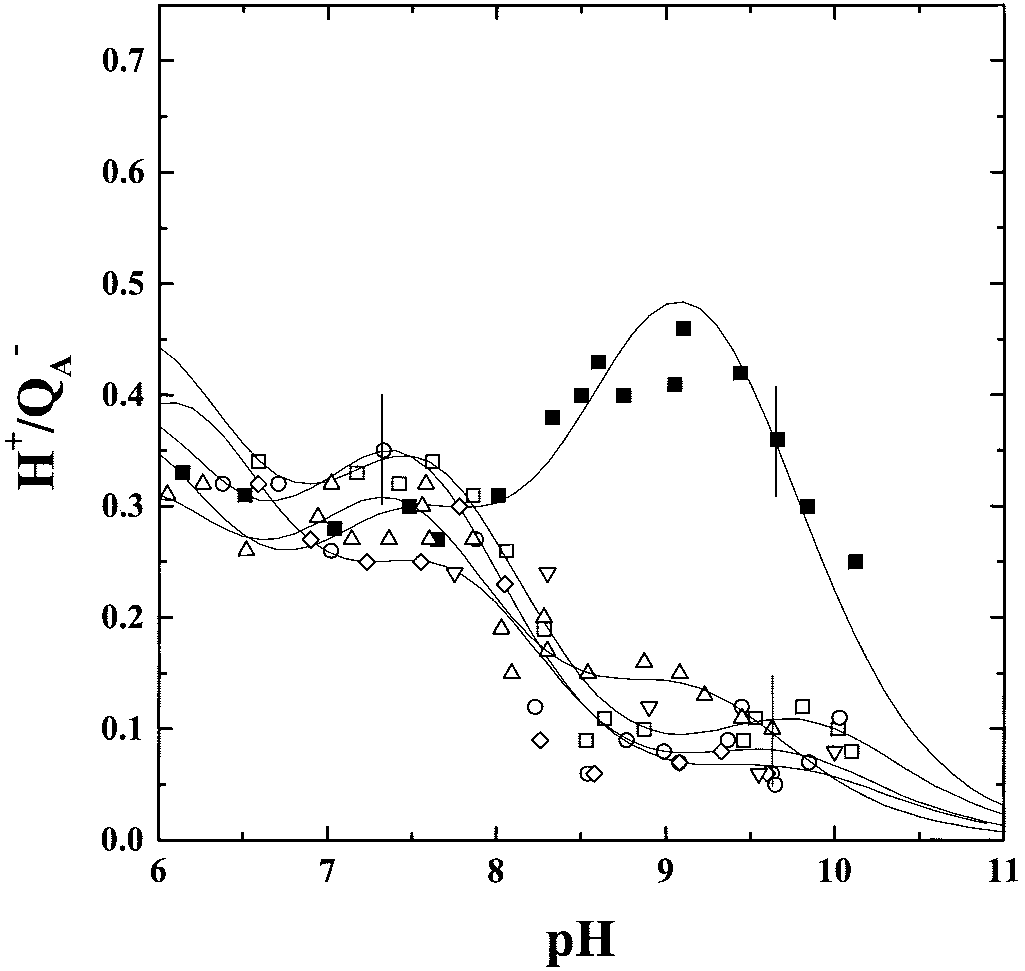
 pH dependence of the stoichiometries of proton uptake by the PQϪ
pH dependence of the stoichiometries of proton uptake by the PQϪ
state in RCs of the WT (■), the L209PE (ᮀ), the L209PT (ƒ), the L209PY(E), the
state in RCs of the WT (■), the L209PE (ᮀ), the L209PT (ƒ), the L209PY(E), the
L209PW (छ), and the L209PF (‚) mutants. Conditions: Ϸ2 M RCs, 0.03%
L209PW (छ), and the L209PF (‚) mutants. Same conditions as in Fig. 2, except:
Triton X-100, 100 M ferrocene, 300 M ferrocyanide, 50 mM NaCl, 100 M
60 M ubiquinone-6 (UQ6) and no terbutryn present. The error bars reflect the
terbutryn, 20 M dye (bromocresol purple, o-cresol red, or o-cresol-
respective experimental error of each set of measurements.
pH dependence of the stoichiometries of proton uptake by the PQϪ
pH dependence of the stoichiometries of proton uptake by the PQϪ
state in RCs of the WT (■), the L209PE (ᮀ), the L209PT (ƒ), the L209PY(E), the
state in RCs of the WT (■), the L209PE (ᮀ), the L209PT (ƒ), the L209PY(E), the
L209PW (छ), and the L209PF (‚) mutants. Conditions: Ϸ2 M RCs, 0.03%
L209PW (छ), and the L209PF (‚) mutants. Same conditions as in Fig. 2, except:
Triton X-100, 100 M ferrocene, 300 M ferrocyanide, 50 mM NaCl, 100 M
60 M ubiquinone-6 (UQ6) and no terbutryn present. The error bars reflect the
terbutryn, 20 M dye (bromocresol purple, o-cresol red, or o-cresol-
respective experimental error of each set of measurements.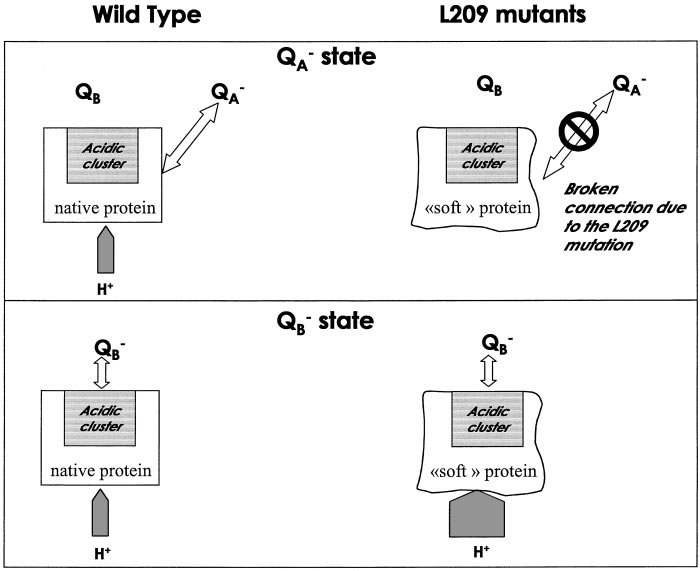 Scheme representing the response of the protein to the formation of QϪ
A and QB in the WT and in the L209P mutants. In the WT, the QA state triggers
the uptake of substoichiometric protons by the cluster from the outside of the protein. In the case of the L209 mutants, the absence of L209Pro softens the protein,altering the connecting relays between the QϪ
A and the QB environment. In the QB state, in the mutants, a substantial additional amount of protons is taken from
the bulk as a consequence of the disorganization of the hydrogen bond network.
Scheme representing the response of the protein to the formation of QϪ
A and QB in the WT and in the L209P mutants. In the WT, the QA state triggers
the uptake of substoichiometric protons by the cluster from the outside of the protein. In the case of the L209 mutants, the absence of L209Pro softens the protein,altering the connecting relays between the QϪ
A and the QB environment. In the QB state, in the mutants, a substantial additional amount of protons is taken from
the bulk as a consequence of the disorganization of the hydrogen bond network.