Tadalafil entfaltet seine Wirkung über eine selektive Hemmung der PDE5, wodurch die Konzentration von cGMP im glatten Muskelgewebe stabil bleibt. Diese biochemische Modulation resultiert in einer langanhaltenden Relaxation der Gefäßwände. Der Wirkstoff wird nach oraler Einnahme effizient resorbiert, mit einer Bioverfügbarkeit von rund 80 %. Seine Halbwertszeit von bis zu 36 Stunden ist innerhalb dieser Substanzklasse außergewöhnlich. Abgebaut wird er in der Leber, hauptsächlich durch CYP3A4, mit anschließender biliärer Exkretion. Typische unerwünschte Wirkungen entstehen durch eine verstärkte Vasodilatation, etwa Kopfschmerzen oder Flush. Pharmakologisch wird cialis generika vor allem durch die verlängerte Wirkungsdauer charakterisiert.
Mk-801 improves retention in aged rats: implications for altered neural plasticity in age-related memory deficits
Neurobiology of Learning and Memory 71, 194 –206 (1999) Article ID nlme.1998.3864, available online at http://www.idealibrary.com on
Implications for Altered Neural Plasticity
Department of Pharmacology, College of Medicine, University of Kentucky,
Alterations in N-methyl-D-aspartate receptor (NMDAR)-dependent synaptic plas-
ticity, characteristic of aged rodents, may contribute to impaired memory withadvanced age. The purpose of the current research was to examine whetherNMDARs contribute to rapid forgetting on a spatial memory task. Aged (22–24months) and adult (3– 6 months) male Fischer 344 rats received 18 training trials,over a period of 3 to 4 h, on the spatial version of the Morris water maze. Immedi-ately after training, a standard free-swim probe trial was administered to assess theacquisition of spatial bias, which was determined by the percent of time spent in thegoal quadrant and the number of platform crossings. Rats then received injectionsof the noncompetitive NMDAR antagonist, (ϩ)-10,11-dihydro-5methyl-5H-dibenzo-(a,b)cycloheptene-5,10 imine (MK-801, 0.05 mg/kg, i.p.), or a vehicle injection ofequal volume. Approximately 24 h later, rats were administered a second free-swimprobe trial to assess retention of spatial bias. All age/drug groups exhibited a spatialbias on the acquisition probe, with adults generally outperforming the aged rats. Onthe retention probe, this spatial bias continued to be shown by adult rats, regardlessof treatment. For the aged group, in contrast, only MK-801-injected rats maintaineda spatial bias on the retention probe, suggesting that NMDAR activity may beinvolved in rapid forgetting during aging. Because blockade of NMDARs also mayimpair new learning, which may, in turn, protect previously stored information fromretroactive interference, rats in a second experiment received post-training injec-tions of scopolamine (0.05 mg/kg), a compound known to inhibit learning. However,scopolamine did not enhance retention in the aged group, consistent with thehypothesis that MK-801 influenced memory in aged rats through its actions onNMDAR-dependent synaptic plasticity.
Aging and age-related neurodegenerative diseases are commonly associated
with a decline in cognitive function. Of particular sensitivity to the aging
This research was supported by the Commonwealth of Virginia Alzheimer’s and Related Dis-
eases Research Award to T.C.F. and a Glenn Foundation/AFAR award to C.M.N. Also supportedby National Institute of Health Grant AG-14979.
Address correspondence and reprint requests to Dr. Thomas Foster, Department of Psychology,
102 Gilmer Hall, University of Virginia, Charlottesville, Virginia 22903-2477. Fax: (804) 982-4685. Alternative mailing address: Dr. Thomas C. Foster, Department of Pharmacology, College ofMedicine MS-305, University of Kentucky, Lexington, Kentucky 40536. Fax (606) 323–1981.
1074-7427/99 $30.00Copyright 1999 by Academic PressAll rights of reproduction in any form reserved.
process are behaviors that depend on the integrity of the hippocampus, such asspatial navigation (Geinisman, deToledo-Morrell, Morrell, & Heller, 1995). Deficits in spatial cognition are consistently observed in aged rodents perform-ing the Morris swim task (Diana, Domenici, de Carolis, Loizzo, & Sagratella,1995; Rasmussen, Schliemann, Sorensen, Zimmer, & West, 1996; Foster,Barnes, Rao, & McNaughton, 1991; Gage, Dunnett, & Bjorklund, 1984; Mabry,McCarty, Gold, & Foster, 1996; Rapp, Rosenberg, & Gallagher, 1987). Ageneral finding is that aged individuals exhibit more rapid forgetting over anextended period of time relative to adults. Although the mechanisms under-lying this diminished retention remain unknown, increasing emphasis hasbeen placed on alterations in hippocampal physiology, including neural plas-ticity (Geinisman et al., 1995; Gallagher, Landfield, McEwen, Meaney, Rapp,Sapolsky, & West, 1996).
(NMDAR) are crucial for the induction of several forms of synaptic plasticity,such as long-term potentiation (LTP) and long-term depression (LTD) (forreview, see Bear & Malenka, 1994; Foster & Norris, 1997). During aging,NMDAR-dependent synaptic plasticity undergoes specific alterations. In par-ticular, induction of LTP using threshold stimulation parameters is dimin-ished (Deupree, Bradley, & Turner, 1993; Moore, Browning, & Rose, 1993),and the decay of LTP over a period of days is accelerated in memory-impairedaged rats (Barnes & McNaughton, 1985; deToledo-Morrell, Geinisman, &Morrell, 1988). In contrast to LTP, susceptibility to the induction of LTD andthe reversal of LTP is augmented during aging (Norris, Korol, & Foster, 1996),providing a rare example of increased synaptic plasticity with advanced age. The importance of this finding may be considerable, since theoretical reportsand empirical data indicate that synaptic function is regulated by the inter-action of LTP and LTD mechanisms (Hrabetva & Saktor, 1996; Mayford,Wang, Kandel, & O’Dell, 1995; Oliet, Malenka, & Nicoll, 1996; Oliet, Malenka,& Nicoll, 1997). Consequently, it has been suggested that enhanced LTDduring aging may underlie the aforementioned age-related deficits in LTP,making synaptic depression a potential candidate mechanism for increasedforgetting (Foster & Norris, 1997; Norris et al., 1996).
Because LTD and LTP-reversal in the aged rat are inhibited by antagonism
of the NMDAR (Norris et al., 1996), the current research was undertaken totest the paradoxical hypothesis that NMDAR blockers, which disrupt theacquisition of spatial information when given before training, should facilitateretention of a spatial task when given after training. However, in addition toblocking synaptic plasticity it was reasonable to suspect that MK-801 couldaffect memory through other processes, such as the inhibition of new learningand retroactive interference. Thus, in a second experiment, aged and adult ratsreceived post-training injections of another compound known to inhibit learn-ing (i.e., scopolamine). The findings showed that aged rats which receivedinjections of the NMDAR antagonist MK-801 immediately following trainingon the spatial version of the Morris water maze continued to exhibit significantretention of the spatial task over a 24-h period. In contrast, aged rats thatreceived post-training injections of scopolamine or vehicle did not exhibitsignificant retention over 24 h. The results are consistent with the hypothesisthat age-related changes in NMDAR-dependent synaptic plasticity are in-volved in impaired retention during aging.
Subjects were 23 adult (3– 6 mos) and thirty-three aged (20 –24
mos) male Fischer-344 rats, obtained from the National Institute of Aging’scolony at Harlan. Animals were individually caged, maintained on a 12:12 hlight– dark cycle, and had access to rat chow and water ad libitum.
The water maze was a black circular pool (164 cm in diameter,
45 cm in height) located in a well lit room and filled with water heated to 27°C. The subject’s movement in the pool was recorded on videotape and analyzedoff-line using a Columbus Instruments tracking system. A black platform (12cm in diameter), located in the center of one of four quadrants of the pool,allowed rats to escape the water.
Animals received intraperitoneal (i.p.) injections of either (ϩ)-
10,11-dihydro-5methyl-5H-dibenzo(a,b)cycloheptene-5,10 imine (MK-801, Re-search Biochemicals International) (0.05 mg/kg, dissolved in H O) or vehicle
alone. Final volume for both MK-801 and vehicle injections was 1 mL/kg. Thisdose was chosen because previous research indicates that ϳ0.05 mg/kg MK-801 impairs the acquisition of spatial information without eliciting motordisturbances over a 24-h period (Jerram, Smith, & Darlington, 1996; McLambet al., 1990; Parada-Turska & Turski, 1990; Whishaw & Auer, 1989; Ylinen,Pitkanen, Sirvio, Hartikainen, Sivenius, Koivisto, & Riekkinen, 1995). Ratswere randomly assigned to each drug condition.
Visual contrast between the escape platform
and the surrounding room was increased by wrapping the escape platformwith white tape, elevating it 1.5 cm above the water’s surface, and encirclingthe pool with black curtains. Prior to cue discrimination training, rats wereplaced in the water and permitted to mount the escape platform from threedifferent locations. Following the third mount, rats remained on the escapeplatform for 60 s before being returned to their home cages to await the firsttraining trial (ϳ20 min). Training consisted of six blocks of three trials, with anintertrial interval of 30 s and an interblock interval of 20 –30 min. Ratsremained on the platform between trials and in home cages (under a heatlamp) between blocks. On each trial, the rat was placed in the water (facing thewall) at one of eight equally spaced locations along the perimeter of the pool. Starting and platform locations were randomized across trials. Rats werepermitted 60 s to navigate to the escape platform. If the platform was notlocated within the allotted time, rats were gently guided to the platform wherethey remained until the next trial. Path length traveled to escape was used tomeasure performance on each trial. Animals that could not consistently nav-igate to the platform were assumed to have sensory-motor, or motivationaldeficits and were eliminated from the study. Three days following the cuediscrimination task, the remaining rats received spatial training.
In the spatial task, the escape platform was all black and
located just beneath the water’s surface. The platform remained in the samequadrant across trials and the curtains surrounding the pool were removed,revealing a number of extra-maze cues throughout the room (computer, ladder,
cabinets, wall posters, lamps). Starting locations were randomized acrosstrials. Training consisted of six blocks of three trials (60 s/trial) with anintertrial interval of 30 s and an interblock interval of 20 –30 min. Path lengthtraveled to escape was used to measure performance on each trial. Acquisition and retention testing.
After the last spatial training block (ϳ20
min), a probe trial (acquisition probe) was delivered. In probe trials, theplatform was removed from the pool allowing the rats to swim freely for 60 s. After the acquisition probe, the platform was reintroduced to the pool (in thesame location as during training) and rats received a “refresher” block of threetrials. Rats were then injected with either MK-801 or vehicle and returned totheir home cages where they remained for approximately 24 h until delivery ofa second probe trial (retention probe). Percent time spent searching eachquadrant and the number of platform crossings (number of times the animalcrosses the area where the platform was located) was calculated for each probetrial.
For training on cue and spatial discrimination tasks,
path length traveled by each animal was averaged across the three trialswithin each block. These block means were subjected to repeated-measuresANOVA to detect effects of training and age. Percent time spent in the goalquadrant and the number of platform crossings for each of the probe trialswere also analyzed using repeated measures ANOVA. For each of the fourexperimental groups (aged vehicle, aged MK-801, adult vehicle, adult MK-801), Student’s t-tests were used to determine whether the percent time spentin the goal quadrant differed significantly from chance (i.e., 25%). Significancefor all statistical tests was set at p Ͻ .05.
Fifteen aged rats were unable to complete swim train-
ing in the cue discrimination task and were not included in the analyses. Theresults of the cue discrimination task are illustrated in Fig. 1A. Significanteffects of age [F(1, 175) ϭ 17.725, p Ͻ 0.001], training [F(5, 175) ϭ 5,195, p Ͻ0.0001], and an age ϫ training interaction [F(5, 175) ϭ 2.4, p Ͻ 0.05] werefound. Although both age groups exhibited a decrease in path length from thefirst to the last training block [aged, F(5, 70) ϭ 3.4, p Ͻ 0.01, n ϭ 16]; adult,F(5, 105) ϭ 11.22, p Ͻ 0.0001, n ϭ 23], adults showed a greater reduction inpath length by the end of training.
For spatial discrimination training, significant ef-
fects of age [F(1, 210) ϭ 9.791, p Ͻ 0.01] and training [F(6, 210) ϭ 35.159, p Ͻ0.0001] were revealed (Fig 1B). Again, even though both age groups exhibitedlearning [aged, [F(6, 84) ϭ 14.119, p Ͻ 0.0001]; adult F(6, 126) ϭ 23.574, p Ͻ0.0001], path length for adults was significantly shorter than that for the agedgroup. Acquisition and retention testing.
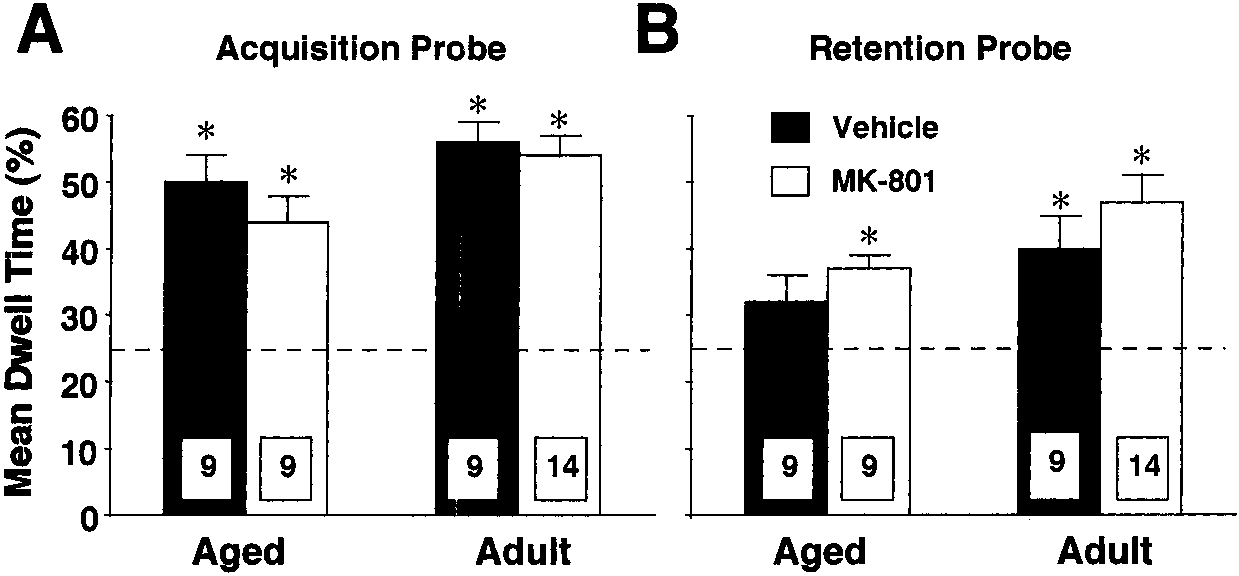
tion probes for each age/drug group is shown in Fig. 2. Overall, adults spentmore time searching the goal quadrant on both probe trials. Follow-up ANO-VAs within each age group indicated a trial ϫ drug interaction only for agedrats [F(1, 14) ϭ 4.616, p Ͻ 0.05], attributable to better performance by MK-
Effect of age on acquisition of cue (A) and spatial (B) discrimination in Experiment 1.
Mean path length for aged (filled circles) and adult rats (open circles) is plotted on the y axis as afunction of training block. Error bars equal SEM.
801-treated rats on the retention probe relative to age-matched vehicle-treatedcontrols. Further inspection using Student’s t-tests indicated that, for theacquisition probe, all groups exhibited differential search strategies (i.e., per-cent of time in goal quadrant was greater than 25%) [aged vehicle, t (8) ϭ 6.08,p Ͻ 0.001; aged MK-801, t (8) ϭ 5.209, p Ͻ 0.001; adult vehicle, t (13) ϭ 8.153,p Ͻ 0.0001; adult MK-801, t (8) ϭ 8.739, p Ͻ 0.0001] (Fig 2A). Conversely,during the retention probe, this differential search strategy was maintained byall groups except the aged vehicle-treated group [aged MK-801, t (8) ϭ 5.875,p Ͻ 0.001, adult vehicle, t (13) ϭ 2.9, p Ͻ 0.02; adult MK-801, t (8) ϭ 5.563, p Ͻ0.001] (Fig 2B).
The improved retention performance of aged animals, treated with MK-801,
was confirmed by analyses of platform crossings (Table 1). Again, adultsexhibited greater performance on both probe trials [F(1, 34) ϭ 11.087, p Ͻ0.001] and an overall reduction in the number of platform crossings was
Post-training administration of MK-801 enhances retention of place learning. Mean
dwell time in the goal quadrant (% of total swim time) for acquisition (A) and retention (B) probetrials is plotted on the y axis. Animals receiving vehicle or MK-801 are represented by filled andopen columns, respectively. Dashed line indicates the percent dwell time expected from chancealone (i.e., 25%). Asterisks denote a significant difference from chance. Note that in the retentionprobe (B), all groups but the aged-vehicle treated rats performed above chance levels. n for eachage/drug group is provided in the appropriate column. Bars equal SEM. Number of Goal Crossings in Acquisition and Retention Probes for Experiment 1 Note. Values are means ϮSEM. Asterisks next to retention probe values indicate significant
reductions from respective acquisition probe values. p Ͻ .05.
observed across the 24 h interval. Interestingly, the age ϫ trial interactionreached significance [F(1, 34) ϭ 10.235, p Ͻ 0.01], and the age ϫ trial ϫ druginteraction approached significance [F(1, 34) ϭ 3.28, p ϭ 0.07]. Follow-upANOVAs within each age group demonstrated a trial ϫ drug interaction onlyfor aged rats [F(1, 17) ϭ 6.274, p Ͻ 0.02]. These results were due to the factthat, in contrast to all other age/drug groups, MK-801-treated aged rats did notexhibit a substantial reduction in the number of platform crossings from theacquisition to the retention probe trial [aged vehicle, F(1, 17) ϭ 14.4, p Ͻ 0.01;adult vehicle, F(1, 25) ϭ 42.26, p Ͻ 0.0001; adult MK-801, F(1, 25) ϭ 14.09, p Ͻ0.01]. Together, the evidence suggests that MK-801 improves retention in anage-dependent manner.
The findings of the previous experiment indicate that post-training injec-
tions of MK-801 enhance the retention of spatial discrimination in aged rats. Although the results are consistent with the idea that MK-801 improvesmemory via blockade of synaptic plasticity, it should be noted that this druginfluences a number of other neural processes that may also contribute tomemory function. For instance, NMDAR antagonists impair the acquisition ofnew information (e.g., Caramanos & Shapiro, 1994; Morris, 1988; Morris,Anderson, Lynch, & Baudry, 1986; Robinson, Crooks, Shinkman, & Gallagher,1989). As such, MK-801 may protect previously stored information from ret-roactive interference.
To address this alternative possibility in Experiment 2, rats were injected
with the cholinergic antagonist scopolamine following spatial discriminationtraining. Similar to MK-801, scopolamine impairs the acquisition of spatialinformation (e.g. Markowska, Olton, & Givens, 1995). Unlike NMDAR antag-onists, however, cholinergic antagonists have little or no effect on the inductionof LTD (Bear & Abraham, 1996).
All aspects of this experiment were identical to those in Experiment 1. The
only exception being that rats (seven adult and five aged) were injected with
scopolamine hydrobromide (0.05 mg/kg, i.p.) following spatial discriminationtraining. Seven adult and five aged rats were used in this experiment.
Figure 3A illustrates performance on cue discrimina-
tion training. A significant effect of training was found [F(5, 50) ϭ 4.605, p Ͻ0.01], indicating that, overall, path length decreased across blocks.
For spatial discrimination training, significant ef-
fects of age [F(1, 60) ϭ 5.98, p Ͻ 0.05] and training [F(6, 60) ϭ 18.195, p Ͻ0.0001] were revealed (Fig 3B). Similar to Experiment 1, both age groupsexhibited learning [aged, [F(6, 34) ϭ 7.6, p Ͻ 0.0001]; adult F(6, 48) ϭ 11.69,p Ͻ 0.0001], however, path length for adults was significantly shorter thanthat for the aged group. Acquisition and retention testing.
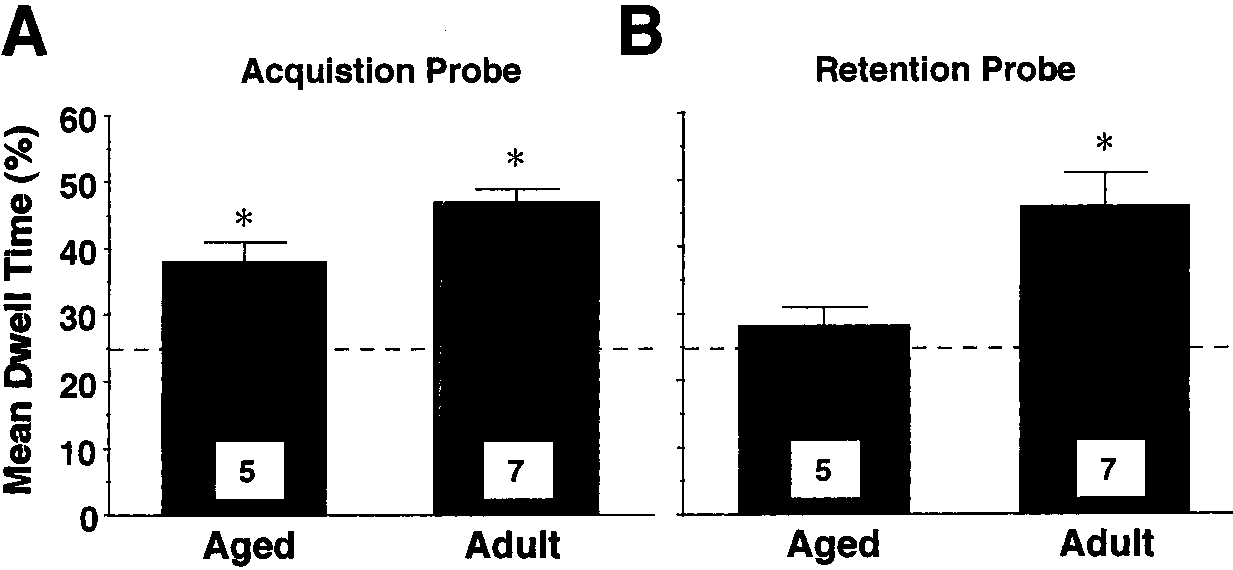
tion probes for each age group is shown in Fig. 4. A repeated measures ANOVArevealed a significant effect of age [F(1, 10) ϭ 12.29, p Ͻ 0.01], in which adultsspent significantly more time than aged rats in the goal quadrant on bothprobe trials. Student’s t-tests comparing the percent time spent in the goalquadrant to chance indicated that, for the acquisition probe, both groupsexhibited differential search strategies [aged, t (4) ϭ 4.24, p Ͻ 0.02; adult, t (6)
ϭ 9.323, p Ͻ 0.0001] (Fig. 4A). During the retention probe, however, thisdifferential search strategy was maintained only for the adult group [t (6) ϭ3.89, p Ͻ 0.01] (Fig. 4B).
Analysis of the number of platform crossings for the acquisition and reten-
tion probes (Table 2) revealed significant effects of age [F(1, 10) ϭ 17.55, p Ͻ0.01] and trial [F(1, 10) ϭ 13.77, p Ͻ 0.01]. Overall, adults crossed the escapeplatform more than aged rats on both probe trials and the number of crossingsdecreased on the retention probe trial for both age groups [aged, F(1, 9) ϭ 7.11,p Ͻ 0.05; adult [F(1, 13) ϭ 8.23, p Ͻ 0.05]. Thus, aged rats injected withscopolamine failed to exhibit significant retention of spatial information, sim-ilar to vehicle-treated aged rats in Experiment 1.
The main conclusion of this study is that the NMDAR antagonist, MK-801,
enhances retention in aged rats when administered after training. Similarmemory-enhancing effects of MK-801 have been reported for young animalsusing other hippocampal-dependent behavioral assays (Mondadori, Weisk-rantz, Buerki, Petschke, & Fagg, 1989). However, other reports have indicatedno effect, or impaired retention of a water maze task following post-traininginjections of MK-801 (Mondadori et al., 1989; Packard & Teather, 1997). Thecurrent study had distinct advantages for examining the acquisition andretention of spatial information. First, in the current report, rats underwent 18training trials over the course of 2–3 h, whereas daily training sessions in theother studies consisted of only one to eight trials delivered over the course ofminutes. Thus, the more extensive training endured by rats in the currentresearch may have permitted a greater level of acquisition prior to drugdelivery. Second, spatial discrimination in the current report was determined
Effect of age on acquisition of cue (A) and spatial (B) discrimination in Experiment 2.
Mean path length for aged (filled circles) and adult rats (open circles) is plotted on the y axis as afunction of training block. Error bars equal SEM.
using “free-swim” probe trials, which facilitates the distinction between spatialand nonspatial strategies. In contrast, the previous studies used only escapelatency measures. While a decrease in latency is indicative of learning, it isunclear from this measure alone whether a spatial or non-spatial strategy isemployed to navigate to the platform (Gallagher, Burwell, & Burchinal, 1993). This problem may be substantial if MK-801 influences spatial and nonspatialmemory in different ways.
Although both age groups exhibited learning on cue and spatial tasks, age
differences in path length were observed in both Experiments 1 and 2. Thisdifference may be related to the age of the animals tested. Our previous workcomparing 9- and 24-mos old animals indicated only small differences in pathlength using a similar training paradigm (Foster et al., 1991). Perhaps moreimportantly, the pool used in the current report was considerably larger thanthat used in our previous study (164 cm vs 120 cm), which undoubtedly
Effects of scopolamine on the retention of place learning. Mean dwell time in the goal
quadrant (% of total swim time) for acquisition (A) and retention (B) probe trials is plotted on they axis. Dashed line indicates the percent dwell time expected from chance alone (i.e., 25%). Asterisks denote a significant difference from chance. Note that in the retention probe (B), onlyadults performed above chance levels. n for each age/drug group is provided in the appropriatecolumn. Bars equal SEM. Number of Goal Crossings in Acquisition and Retention Probes for Experiment 2 Note. Values are means ϮSEM. Asterisks next to retention probe values indicate significant
reductions from respective acquisition probe values. p Ͻ 0.05.
lengthened the amount of swimming, especially during the early phases oftraining. In fact, path length for each age group on the first two training blockswas more than double that reported by Foster et al. (1991). Thus, the highelimination level of aged rats following the cue discrimination task, and theinability of the remaining aged rats to achieve adult-like levels of performancemay be attributable to a fatigue effect, rather than a learning deficit.
There were at least two indications that MK-801’s effects on retention
were more pronounced for aged animals. First, while all MK-801-treatedrats spent more time in the goal quadrant on the retention probe thancontrol rats, only the aged control group failed to search the goal quadrantabove chance levels during retention testing. Thus, in contrast to adults,the retention differences between MK-801-treated and vehicle-treated agedrats may have been underestimated, since search time for the aged controlgroup would be expected to drop no lower than 25%. The finding thatvehicle-treated aged rats failed to exhibit a differential search strategy onthe retention probe trial is similar to previous reports that have observedrapid forgetting in aged animals (e.g. Barnes & McNaughton, 1985; Dianaet al, 1995; Rasmussen et al, 1996; Foster et al., 1991; Gage et al., 1984;Gold, McGaugh, Hankins, Rose, & Vasquez, 1981; Linder, Balch, & Van-derMaelen, 1992; Mabry et al., 1996; Rapp et al., 1987; Winocur, 1988). Secondly, when the number of platform crossings was examined for bothprobe trials (Tables 1 and 2), each group except the aged MK-801 ratsexhibited a drop in the number of crossings from the acquisition probe tothe retention probe, indicating that aged rats injected with MK-801 contin-ued to focus their search to the location that had contained the platform.
The benefits of MK-801 in facilitating retention may relate to altered syn-
aptic plasticity during aging. Specifically, NMDAR blockade may inhibit LTD-like processes which are enhanced in the aged rat. Functional modifications inhippocampal synaptic transmission properties provide the best correlates forcognitive decline during aging, and as such, may underlie rapid forgetting ontasks that require an intact hippocampus (Barnes, Treves, Rao, & Shen, 1994;Gallagher et al., 1996; Geinisman et al., 1995). It has been suggested that theincreased susceptibility to LTD in aged rats may contribute to reduced synap-tic strength and accelerated LTP decay (Foster & Norris, 1997). Thus, a shiftin plasticity processes, in favor of LTD, may provide a mechanistic basis forrapid forgetting in the aged animal (Foster & Norris, 1997).
This argument predicts that NMDAR antagonists, which inhibit LTD induc-
tion, should facilitate retention of spatial information when given after acqui-sition. The results of the current study were consistent with this prediction. Alternatively, NMDAR blockade may influence other processes that couldcontribute to forgetting, such as retroactive interference, or the modulation ofneural (i.e. theta) activity (Pitkanen et al., 1995; however also see Whishawand Auer, 1989). However, in the present study, aged rats injected withscopolamine at a dose shown to inhibit learning and theta activity (Markowskaet al., 1995), failed to exhibit significant retention of spatial information. Theresults are, at the least, consistent with the idea that MK-801 differentiallyinfluenced memory in aged and adult rats by interacting with age-dependentchanges in synaptic plasticity. Some caution in interpreting this latter findingis warranted, however, since spatial bias in the aged group on the acquisitionprobe, although significant, was not striking. The parametric space availablefor determining a statistical effect of scopolamine for aged rats therefore mayhave been relatively limited. It will be important for future research to deter-mine if the effects of MK-801 on retention in aged rats are specific to its actionson hippocampal synapses and also to specify whether MK-801 inhibits LTD inaged animals at a behaviorally relevant dose.
The results raise the intriguing possibility that normal learning and mem-
ory reflect the balance and influence of LTP and LTD. As such, the increasedrate of forgetting in older animals may result from an activity-dependentmechanism analogous to LTD. Similar to aged rats, adult rats can also exhibitNMDAR-dependent synaptic depression, albeit the susceptibility to LTD-in-duction is reduced in young adults (Foster and Norris, 1997; Norris, et al.,1996). If processes which are mechanistically similar to LTD underlie forget-ting, then one would expect that NMDAR blockade should improve retention inyounger animals as well. While adult rats injected with MK-801 generallyspent more time in the goal quadrant on the retention probe than age-matchedcontrol rats (Figure 2), a significant effect of drug was not observed. Becausethe 24 hr interval between the two free-swim probes does not appear tochallenge the retention ability of adults, (i.e. all adults perform well on theretention probe), it is plausible that our retention task lacked the sensitivity toreveal any effects of MK-801 on memory in the adult group.
Induction of LTD is a Ca2ϩ-dependent process. Dysregulation of Ca2ϩ is a
common observation in neurons from aged mammals. A contributing source toaltered regulation of neuronal Ca2ϩ is an increase in L-type voltage-dependentCa2ϩ channels (Campbell, Hao, Thibault, Blalock, & Landfield, 1996; Thibault& Landfield, 1996). The fact that L-channel blockade ameliorates age-relatedchanges in neurophysiology (Moyer & Disterhoft, 1994; Moyer, Thompson,Black, & Disterhoft, 1992), synaptic plasticity (Norris, Halpain, & Foster,1998) and cognition (Deyo, Straube, & Disterhoft, 1989; Ingram, Joseph,Spangler, Roberts, Hengemihle, & Fanelli; Levere & Walker, 1992; Kowalska& Disterhoft, 1994; Straube, Deyo, Moyer, & Disterhoft, 1990; Soloman, Wood,Groccia-Ellison, Yang, Fanelli, & Mervis, 1995) indicates a potentially impor-tant link between Ca2ϩ-dependent processes, such as synaptic plasticity, andmemory. Together, the results may provide a useful theoretical framework forexploring the neurobiological basis of cognitive decline during aging.
Barnes, C. A., & McNaughton, B. L. (1985). An age comparison of the rates ofacquisition and
forgetting of spatial information in relation to long-term enhancement of hippocampal syn- apses. Behavioral Neuroscience, 99, 1040 –1048.
Barnes, C. A., Rao, G., Foster, T. C., & McNaughton, B. L. (1992). Region-specific age effects on
AMPA sensitivity: Electrophysiological evidence for loss of synaptic contacts in hippocampal field CA1. Hippocampus, 2, 457– 468.
Barnes, C. A., Rao, G., & McNaughton, B. L. (1996). Functional integrity of NMDA-dependent LTP
induction mechanisms across the lifespan of F-344 rats. Learning and Memory, 3, 124 –137.
Barnes, C. A., Treves, A., Rao, G., & Shen, J. (1994). Electrophysiological markers of cognitive
aging: region specificity and computational consequences. Seminars in Neuroscience, 6, 359 – 367.
Bear, M. F., & Malenka, R. C. (1994). Synaptic plasticity: LTP and LTD. Current Opinion inNeurobiology, 4, 389 –399.
Campbell, L. W., Hao, S. Y., Thibault, O., Blalock, E. M., & Landfield, P. W. (1996). Aging changes
in voltage-gated calcium currents in hippocampal CA1 neurons. The Journal of Neuroscience, 16, 6286 – 6295.
Caramanos, Z., & Shapiro, M. L. (1994). Spatial memory and N-methyl-D-aspartate receptor
antagonists APV and MK-801: Memory impairments depend on familiarity with the environ- ment, drug dose, and training duration. Behavioral Neuroscience, 108, 30 – 43.
deToledo-Morrell, L., Geinisman, Y., & Morrell, F. (1988). Age-dependent alterations in hippocam-
pal synaptic plasticity: Relation to memory disorders. Neurobiology of Aging, 9, 581–590.
Deupree, D. L., Bradley, J., & Turner, D. A. (1993). Age-related alterations in potentiation in the
CA1 region in F344 rats. Neurobiology of Aging, 14, 249 –258.
Deyo, R. A., Straube, K. T., & Disterhoft, J. F. (1989). Nimodipine facilitates trace conditioning of
the eyeblink response in aging rabbits. Science, 243, 809 – 811.
Diana, G., Domenici, M. R., de Carolis, A. S., Loizzo, A., & Sagratella, S. (1995). Reduced
hippocampal CA1 Ca2ϩ-induced long-term potentiation is associated with age-dependent impairment of spatial learning. Brain Research, 686, 107–110.
Foster, T. C., Barnes, C. A., Rao, G., & McNaughton, B. L. (1991). Increase in perforant path
quantal size in aged F-344 rats. Neurobiology of Aging, 12, 441– 448.
Foster, T. C., & Norris, C. M. (1997). Age-related changes in Ca2ϩ-dependent processes: relation
to hippocampal synaptic plasticity. Hippocampus, 7, 602– 612.
Gage, F. H., Dunnett, S. B., & Bjorklund, A. (1984). Spatial learning and motor deficits in aged
rats. Neurobiology of Aging, 5, 43– 48.
Gallagher, M., Burwell, R., & Buchinal, M. (1993). Severity of spatial learning impairment in
aging: development of a learning index for performance in the Morris Water Maze. Behavioral Neuroscience, 107, 618 – 626.
Gallagher, M. Landfield, P. W., McEwen, B. Meaney, M. J., Rapp, P. R., Sapolsky, R., & West, M. J.
(1996). Hippocampal neurodegeneration in aging. Science, 274, 484 – 485.
Geinisman, Y., deToledo-Morrell, L., Morrell, F., & Heller, T. E. (1995). Hippocampal markers of
age-related memory dysfunction: Behavioral, electrophysiological and morphological perspec- tives. Progress in Neurobiology, 45, 223–252.
Gold, P. E., McGaugh, J. L., Hankins, L. L, Rose, R. P., & Vasquez, B. J. (1981). Age dependent
changes in retention in rats. Experimental Aging Research, 8, 53–58.
Hrabetva, S., & Sacktor, T. C. (1996). Bidirectional regulation of protein kinase M zeta in the
maintenance of long-term potentiation and long-term depression. The Journal of Neuro- science, 16, 5324 –5333.
Ingram, D. K., Joseph, J. A., Spangler, E. L., Roberts, D., Hengemihle, J., & Fanelli, R. J. (1994).
Chronic nimodipine treatment in aged rats: analysis of motor and cognitive effects and muscarinic-induced striatal dopamine release. Neurobiology of Aging, 15, 55– 61.
Jerram, A. H., Smith, P. F., & Darlington, C. L. (1996). A dose-response analysis of the behavioral
effects of (ϩ)MK-801 in ginea pig: comparison with CPP. Pharmacology Biochemistry and Behavior, 53, 799 – 807.
Kowalska, M., & Disterhoft, J. F. (1994). Relation of nimodipine dose and serum concentration to
learning enhancement in aging rabbits. Experimental Neurology, 127, 159 –166.
Levere, T. E., & Walker, A. (1992). Old age and cognition: enhancement of recent memory in aged
rats by the calcium channel blocker nimodipine. Neurobiology of Aging, 13, 63– 66.
Lindner, M. D., Balch, A. H., & VanderMaelen, C. P. (1992). Short forms of the “reference-”and
“working-memory-” Morris water maze for assessing age-related deficits. Behavioral and Neural Biology, 58, 94 –102.
Mabry, T. R., McCarty, R., Gold, P. E., & Foster, T. C. (1996). Age and stress-history effects on
spatial performance in a swim task in Fischer-344 rats. Neurobiology of Learning and Memory, 66, 1–10.
Markowska, A. L., Olton, D. S., & Givens, B. (1995). Cholinergic manipulations in the medial
septal area: age-related effects on working memory and hippocampal electrophysiology. The Journal of Neuroscience, 15, 2063–2073.
Mayford, M., Wang, J., Kandel, E. R., & O’Dell, T. J. (1995). CaMK-II regulates the frequency-
response function of hippcampal synapses for the production of both LTD and LTP. Cell, 81, 891–904.
McLamb, R. L., Williams, L. R., Nanry, K. P., Wilson, W. A., & Tilson, H. A. (1990). MK-801
impedes the acquisition of a spatial memory task in rats. Pharmacology Biochemistry and Behavior, 37, 41– 45.
Mondadori, C., Weiskrantz, L., Buerki, H., Petschke, F., & Fagg, G. E. (1989). NMDA receptor
antagonists can enhance or impair learning performance in animals. Experimental Brain Research, 75, 449 – 456.
Moore, C. I., Browning, M. D., & Rose, G. M. (1993). Hippocampal plasticity induced by primed
burst, but not long-term potentiation, stimulation is impaired in area CA1 of aged Fischer 344 rats. Hippocampus, 3, 57– 66.
Morris, R. G. M. (1988). Synaptic plasticity and learning: selective impairment of learning in rats
and blockade of long-term potentiation in vivo by the N-methyl-D-aspartate receptor antag- onist AP5. The Journal of Neuroscience, 9, 3040 –3057.
Morris, R. G. M., Anderson, E., Lynch, G. S., & Baudry, M. (1986). Selective impairment of
learning and blockade of long-term potentiation by an N-methyl-D-aspartate receptor antag- onist, AP5. Nature, 319, 774 –776.
Moyer, J. R., & Disterhoft, J. F. (1994). Nimodipine decreases calcium action potentials in rabbit
hippocampal CA1 neurons in an age-dependent and concentration-dependent manner. Hip- pocampus, 4, 11–18.
Moyer, J. R., Thompson, L. T., Black, J. P., & Disterhoft J. F. (1992). Nimodipine increases
excitability of rabbit CA1 pyramidal neurons in an age-and concentration-dependent manner. Journal of Neurophysiology, 68, 2100 –2109.
Norris, C. M., Halpain, S., & Foster, T. C., (1998). Reversal of age-related alterations in synaptic
plasticity by blockade of L-type Ca2ϩ channels. The Journal of Neuroscience, 18, 3171–3179.
Norris, C. M., Korol, D. L., & Foster, T. C. (1996). Increased susceptibility to induction of long-term
depression and long-term potentiation reversal during aging. The Journal of Neuroscience, 16, 5382–5392.
Oliet, S. H. R., Malenka, R. C., & Nicoll, R. A. (1996). Bidirectional control of quantal size by
synaptic activity in the hippocampus. Science, 271, 1294 –1297.
Oliet, S. H. R., Malenka, R. C., & Nicoll, R. A. (1997). Two distinct forms of long-term depression
coexist in CA1 hippocampal pyramidal cells. Neuron, 18, 969 –982.
Packard, M. G., & Teather, L. A. (1997). Posttraining injections of MK-801 produce a time-
dependent impairment of memory in two water maze tasks. Neurobiology of Learning and Memory, 68, 42–50.
Parada-Turska, J., & Turski, W. A. (1990). Excitatory amino acid antagonists and memory: effect
of drugs acting at N-methyl-D-aspartate receptors in learning and memory tasks. Neurophar- macology, 29, 1111–1116.
Pitkanen, M., Sirvia, J., Ylinen, A., Koivisto, E., & Riekkinen, P. (1995). Effects of NMDA receptor
modulation of hippocampal type 2 theta activity in rats. General Pharmacology, 26, 1065– 1070.
Puma, C., Monmaur, V, Sharif, A., & Monmaur, P. (1996). Intraseptal infusion of selective and
competitive receptor agonist NMDA and the antagonist D-2-amino-5-phosphonopentanoic acid: spectral implications for the physostigmine-induced hippocampal theta rhythm in ure- thane-anesthetized rats. Experimental Brain Research, 109, 384 –392.
Rapp, P. R., Rosenberg, R. A., & Gallagher, M. (1987). An evaluation of spatial information
processing in aged rats. Behavioral Neuroscience, 101, 3–12.
Rasmussen, T., Schliemann, T., Sorensen, J. C., Zimmer, J., & West, M. J. (1996). Memory-
impaired aged rats - No loss of principal hippocampal and subicular neurons. Neurobiology of Aging, 17, 143–147.
Robinson, G. S., Crooks, G. B., Shinkman, P. G., & Gallagher, M. (1989). Behavioral effects of
MK-801 mimic deficits associated with hippocampal damage. Psychobiology, 17, 156 –164.
Soloman, P. R., Wood, M. S., Groccia-Ellison, M. E., Yang, B. Y., Fanelli, R. J., & Mervis, R. F.
(1995). Nimodipine facilitates retention of the classically conditioned nictitating membrane response in aged rabbits over long retention intervals. Neurobiology of Aging, 16, 791–796.
Straube, K. T., Deyo, R. A., Moyer, J. R., Jr., & Disterhoft, J. F. (1990). Dietary nimodipine
improves associative learning in aging rabbits. Neurobiology of Aging, 11, 659 – 661.
Thompson, L. T., Disterhoft, J. F., & Deyo, R. A. (1990). Nimodipine enhances spontaneous activity
of hippocampal pyramidal cells in aging rabbits at a dose that facilitates learning. Brain Research, 535, 119 –130.
Whishaw, I. Q., & Auer, R. N. (1989). Immediate and long-lasting effects of MK-801 on motor
activity, spatial navigation in a swimming pool and EEG in the rat. Psychopharmacology, 98, 500 –507.
Winocur, G. (1988). A neuropsychological analysis of memory loss with age. Neurobiology of Aging,9, 487– 494.
Ylinen, A., Pitkanen, M., Sirvio, J., Hartikainen, T., Sivenius, J., Koivisto, E., & Riekkinen, P. J.
(1995). The effects of NMDA receptor antagonists at anticonvulsive doses on the performance of rats in the water maze task. European Journal of Pharmacology, 274, 159 –165.
Animal (2012), 6:5, pp 815–823 & The Animal Consortium 2011Effects of disodium fumarate on ruminal fermentation andmicrobial communities in sheep fed on high-forage dietsY. W. Zhou1, C. S. McSweeney2, J. K. Wang1 and J. X. Liu1-1Institute of Dairy Science, MoE Key Laboratory of Molecular Animal Nutrition, College of Animal Sciences, Zhejiang University, Hangzhou, China; 2CSIRO LivestockIn

 Neurobiology of Learning and Memory 71, 194 –206 (1999)
Neurobiology of Learning and Memory 71, 194 –206 (1999)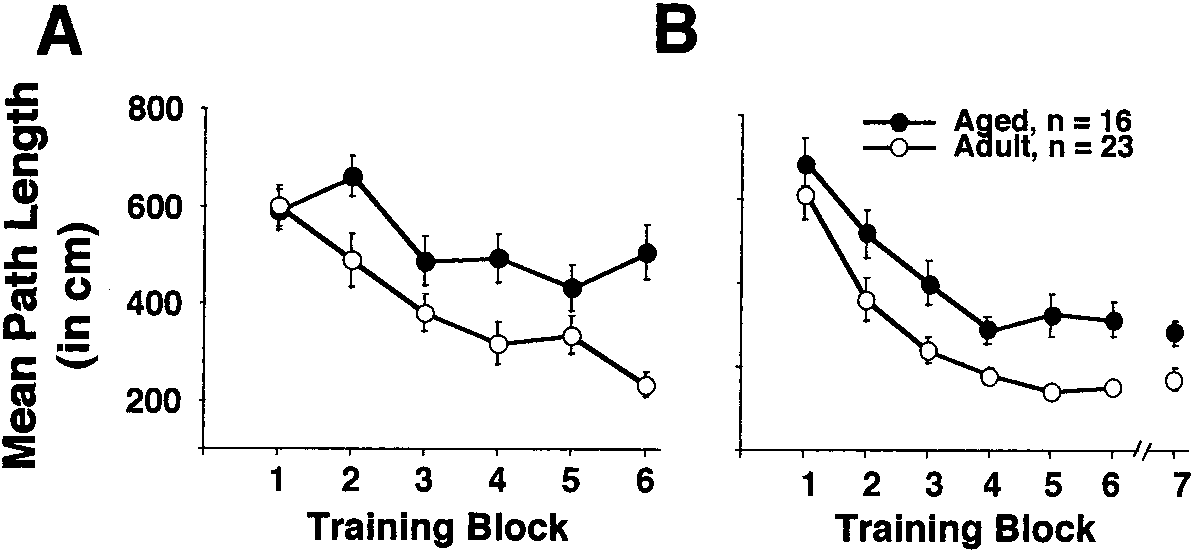
 Effect of age on acquisition of cue (A) and spatial (B) discrimination in Experiment 1.
Effect of age on acquisition of cue (A) and spatial (B) discrimination in Experiment 1.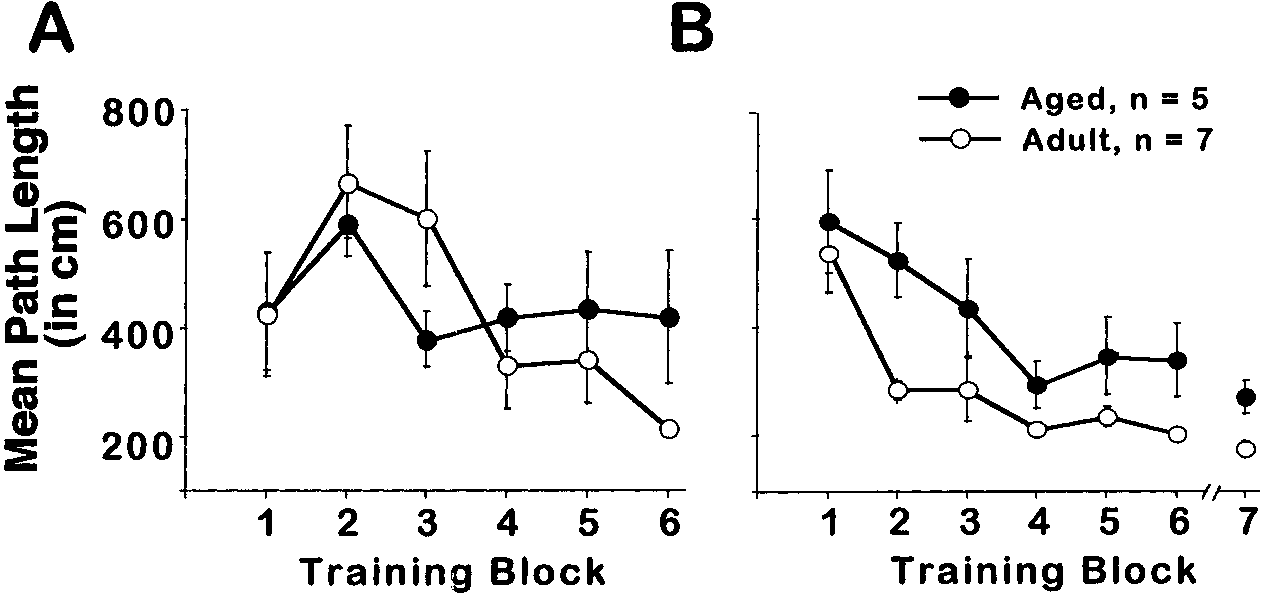
 Effect of age on acquisition of cue (A) and spatial (B) discrimination in Experiment 2.
Effect of age on acquisition of cue (A) and spatial (B) discrimination in Experiment 2.