Tadalafil entfaltet seine Wirkung über eine selektive Hemmung der PDE5, wodurch die Konzentration von cGMP im glatten Muskelgewebe stabil bleibt. Diese biochemische Modulation resultiert in einer langanhaltenden Relaxation der Gefäßwände. Der Wirkstoff wird nach oraler Einnahme effizient resorbiert, mit einer Bioverfügbarkeit von rund 80 %. Seine Halbwertszeit von bis zu 36 Stunden ist innerhalb dieser Substanzklasse außergewöhnlich. Abgebaut wird er in der Leber, hauptsächlich durch CYP3A4, mit anschließender biliärer Exkretion. Typische unerwünschte Wirkungen entstehen durch eine verstärkte Vasodilatation, etwa Kopfschmerzen oder Flush. Pharmakologisch wird cialis generika vor allem durch die verlängerte Wirkungsdauer charakterisiert.
Sb-roscoff.fr
International Journal of Systematic and Evolutionary Microbiology (2004), 54, 41–45
Caminibacter profundus sp. nov., a novelthermophile of Nautiliales ord. nov. within the class‘Epsilonproteobacteria’, isolated from a deep-seahydrothermal vent
M. L. Miroshnichenko,1 S. L’Haridon,2 P. Schumann,3 S. Spring,3E. A. Bonch-Osmolovskaya,1 C. Jeanthon2 and E. Stackebrandt3
Institute of Microbiology, Russian Academy of Sciences, Prospekt 60-letiya Oktyabrya 7/2,
2UMR 6539, Centre National de la Recherche Scientifique and Universite´ de Bretagne
Occidentale, Institut Universitaire Europe´en de la Mer, 29280 Plouzane´, France
3DSMZ – German Collection of Microorganisms and Cell Cultures, Mascheroder Weg 1b,
A novel moderately thermophilic, microaerobic to anaerobic, chemolithoautotrophic bacterium,designated strain CRT, was isolated from a deep-sea hydrothermal vent site at 366N on theMid-Atlantic Ridge. Cells were Gram-negative, non-motile rods. The organism grew at 45–65 6Cand pH 6?5–7?4, with optimum growth at 55 6C and pH 6?9–7?1. The NaCl range for growth was5–50 g l”1 (optimum 30 g l”1). Strain CRT was an obligate chemolithoautotroph, growing withH2 as energy source, sulfur, nitrate or oxygen as electron acceptors and CO2 as carbon source. Hydrogen sulfide and ammonium were the respective products of sulfur and nitrate reduction. The G+C content of the genomic DNA was 32?1 mol%. Based on 16S rRNA gene sequenceanalysis, this organism was most closely related to Caminibacter hydrogeniphilus (94?9 %similarity). On the basis of phenotypic and phylogenetic data, it is proposed that the isolaterepresents a novel species, Caminibacter profundus sp. nov. The type strain is CRT (=DSM15016T=JCM 11957T). The phylogenetic data also correlate well with the significant phenotypicdifferences between the lineage encompassing the genera Nautilia and Caminibacter andother members of the class ‘Epsilonproteobacteria’. The lineage encompassing the genera Nautiliaand Caminibacter is therefore proposed as a new order, Nautiliales ord. nov., represented bya single family, Nautiliaceae fam. nov.
The class ‘Epsilonproteobacteria’ represents a recently recog-
et al., 2003). All the above-mentioned genera, except for
nized line of descent within the Proteobacteria that encom-
Thiovulum and Sulfurospirillum, which thrive in aquatic
pass two families within the single order ‘Campylobacterales’
habitats, have been found associated with animals.
(Garrity & Holt, 2001). The family Campylobacteraceaecontains the genera Campylobacter, Arcobacter, Sulfurospir-
Assessment of microbial diversity using molecular phylo-
illum and Thiovulum, whereas the family ‘Helicobacteraceae’
genetic approaches has revealed that members of the
is formed by the genera Helicobacter and Wolinella. These
‘Epsilonproteobacteria’ dominate various deep-sea hydro-
bacteria are mesophiles adapted to environments that are
thermal habitats such as microbial mats of Loihi Seamount
low in oxygen. Most of them are oxidase-positive micro-
(Moyer et al., 1995), surfaces of invertebrates (Haddad et al.,
aerophiles, but numerous members also grow in the absence
1995; Polz & Cavanaugh, 1995; Cary et al., 1997) and sulfides
of oxygen (Vandamme et al., 1991). Among them, Sulfuro-
from the Mid-Atlantic Ridge (Reysenbach et al., 2000; Corre
spirillum halorespirans and Sulfurospirillum multivorans
et al., 2001) and southern East Pacific Rise (Longnecker &
have been described recently as obligate anaerobes (Luijten
Reysenbach, 2001). Recently, thermophilic representativesof the ‘Epsilonproteobacteria’ have been isolated from tube
Published online ahead of print on 20 June 2003 as DOI 10.1099/
fragments of Alvinella pompejana, an annelid polychaete
endemic to chimney walls of the East Pacific Rise hydro-thermal vents. Both Nautilia lithotrophica and Caminibacter
The GenBank accession number for the 16S rDNA sequence ofCaminibacter profundus strain CRT is AJ535664.
hydrogeniphilus are strictly anaerobic hydrogen-oxidizers
able to grow chemolithoautotrophically with sulfur as
tested on BM medium, with oxygen added to the H2/CO2
electron acceptor (Miroshnichenko et al., 2002; Alain et al.,
mixture (80 : 20, v/v, 200 kPa); the final concentration of
2002). Other organisms that are phylogenetically closely
oxygen varied from 0?25 to 20 %. Carbon source utiliza-
related and phenotypically similar to these species have been
tion was determined using substrates at a concentration of
partially characterized by Campbell et al. (2001). All these
0?05 %; in this case, the headspace was filled with 100 % H2
thermophilic isolates, along with a number of environ-
(atmospheric pressure). Inoculated tubes were incubated at
mental sequences retrieved from hydrothermal systems,
55 uC. The cell density was determined by direct cell count-
form a deep monophyletic unit within the ‘Epsilon-
ing using a light microscope. Gaseous and liquid fermenta-
proteobacteria’. Very recently, many novel phylogenetically
tion products, as well as the products of nitrate reduction,
diverse representatives of the ‘Epsilonproteobacteria’ have
were detected as described previously (Miroshnichenko
been isolated from the hydrothermal fields of the Okinawa
et al., 1994, 2003). Hydrogen sulfide was measured by a
Trough and Central Indian Ridge and partially described
colorimetric method (Tru¨per & Schlegel, 1964). The
(Takai et al., 2003). Here, a second species in the genus
sensitivity of strain CRT to rifampicin, chloramphenicol,
Caminibacter, Caminibacter profundus sp. nov., isolated from
vancomycin, penicillin, streptomycin and tetracycline
a hydrothermal vent of the Mid-Atlantic Ridge, is described.
(Sigma) was tested at a concentration of 100 mg ml21. Determination of the DNA G+C content was performed as
Strain CRT was isolated from material collected using a vent
described elsewhere (Miroshnichenko et al., 2003). DNA
cap at the Rainbow hydrothermal vent field (36u169N;
extraction, PCR amplification of the 16S rRNA gene and
33u549W; 2400 m depth) on the Mid-Atlantic Ridge during
determination of the sequence followed described methods
the Iris cruise in May 2001. An in situ growth chamber or
(Rainey et al., 1996). The 16S rRNA sequences were aligned
vent cap (Reysenbach et al., 2000), designed to concentrate
with published sequences of the DSMZ database using
the micro-organisms discharged by hydrothermal emis-
the ae2 editor (Maidak et al., 1999) and sequences retrieved
sions, was deployed using the hydraulic arm of the remotely
from EMBL. Evolutionary distances were calculated by the
operated vehicle Victor. After incubation in situ for 2 days,
method of Jukes & Cantor (1969). Distance analysis dendro-
the vent cap was closed by the hydraulic arm of the remotely
grams were reconstructed by the neighbour-joining algo-
operated vehicle before transportation to the surface. Once
rithm. Bootstrap analysis was used to evaluate the tree
on board, the vent cap content was immediately transferred
topology by performing 500 resamplings (Felsenstein, 1988).
to 50 ml glass vials and flooded with a sterile solution of3 % (w/v) sea salts (Sigma). The vials were then closed
Enrichment was performed in Bellco tubes filled with 5 ml
tightly with butyl rubber stoppers (Bellco), pressurized with
BM medium. A H2/CO2 mixture (80 : 20, 200 kPa) served as
the energy and carbon source, and elemental sulfur was the
2 (100 kPa), reduced with sodium sulfide and stored at
4 uC until further processing in the laboratory.
electron acceptor. After inoculation of BM medium with0?5 ml material recovered from the vent cap content and
For enrichment, the following basal medium (BM) was used
inner surfaces and incubation of the tubes for 3 days at 55 uC
without shaking, growth of non-motile rods was observed,
accompanied by the formation of hydrogen sulfide. Transfer
2PO4, 0?33; CaCl2.2H2O, 0?33; MgCl2.6H2O, 0?33; NaCl,
25?0; yeast extract, 0?1; trace elements (Balch et al., 1979),
of the enrichment culture into BM medium without yeast
10 ml l21; vitamins (Wolin et al., 1963), 10 ml l21. The
extract did not affect its growth. A pure culture, CRT, was
medium was prepared anaerobically and dispensed into
isolated by serial dilutions in liquid mineral medium. Purity
Bellco tubes; the headspace (25 ml) was filled with H
of the culture was checked by the absence of growth in a
(80 : 20, 200 kPa). No reducing agents were added to the
non-selective glucose- and peptone-containing medium
medium. Elemental sulfur was added to a final concentra-
tion of 10 g l21. The pH of the medium was adjusted with
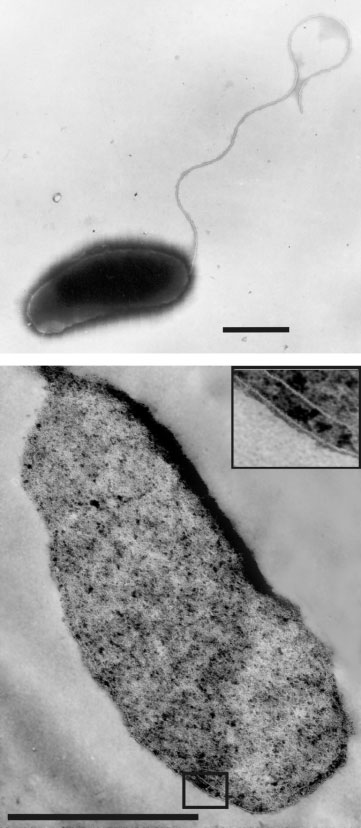
Cells of strain CRT were rod-shaped (approximately 1?2–
2?5 M H2SO4 to 6?8–7?0. When substrates other than mole-
1?560?5 mm) and motile in the exponential phase of
cular hydrogen were tested, the headspace was filled with
growth. One polar flagellum was present on negatively
N2/CO2 (8 : 2, v/v, atmospheric pressure). A pure culture
stained whole-cell preparations (Fig. 1a). Formation of
was obtained on the same basal medium without yeast
spores was not observed. Thin sectioning revealed the
extract using a serial tenfold dilution technique. Morpho-
Gram-negative structure of the cell wall (Fig. 1b).
logy of the novel isolate was examined using an OlympusBX-60 microscope. The ultrastructure of whole cells and
Strain CRT grew anaerobically with molecular hydrogen as
thin sections was studied as described elsewhere (Bonch-
the energy source and elemental sulfur or nitrate as the
Osmolovskaya et al., 1990). For physiological studies, the
electron acceptors. The only product detected during
isolate was grown on BM medium containing MOPS
growth with S0 was H2S. Ammonium was the only product
(10 mM) as a buffer. The pH of the medium was adjusted to
of nitrate reduction. Strain CRT was also able to grow
7?0 with 5 M NaOH before autoclaving. Potential growth
microaerobically at low oxygen concentrations (up to 2 %,
substrates and electron acceptors were added at concentra-
optimal at 0?5 %). With hydrogen, S0 and CO2 as electron
tions of 0?3 and 0?2 % (w/v), respectively. The ability of the
donor, electron acceptor and carbon source, respectively,
isolate to grow microaerobically and/or aerobically was
the isolate grew at 45–65 uC, with optimum growth around
International Journal of Systematic and Evolutionary Microbiology 54
100 mg ml21). It grew in the presence of chloramphenicoland tetracycline (both at 100 mg ml21). The G+C contentof the DNA of isolate CRT was 32?1 mol%.
Comparison of the 16S rRNA gene sequence (1414 bases)with those of members of the domain Bacteria indicatedthat strain CRT belonged to the class ‘Epsilonproteobacteria’and was moderately related to C. hydrogeniphilus (94?9 %similarity) and N. lithotrophica (91?2 % similarity), both ofwhich were isolated from 13uN on the East PacificRise. Strain CRT showed higher sequence similarity(92?3–96?1 %) to a group of clone sequences retrievedfrom material from deep-sea hydrothermal vents on theMid-Atlantic Ridge (VC2.1Bac7, VC2.1Bac17, VC2.1Bac8,
VC2.1Bac30; Reysenbach et al., 2000). Slightly lowersimilarities (91?4–93?7 %) were found to clone sequencesretrieved from South-East Pacific vents (S17sBac14,S17sBac3, S17sBac5; Longnecker & Reysenbach, 2001) andto isolate AM1115 (Alain et al., 2002).
The phylogenetic relatedness of strain CRT to C. hydro-geniphilus is consistent with shared physiological character-istics and the DNA G+C content (Table 1). Both strains aremoderately thermophilic chemolithoautotrophs, growingwith hydrogen as electron donor and elemental sulfur ornitrate as electron acceptors. However, C. hydrogeniphilushas been described as a strictly anaerobic micro-organism,whereas strain CRT is able to grow anaerobically and micro-aerobically at an oxygen concentration of up to 2 %. Theisolate has a narrow pH growth optimum of 6?9–7?1,whereas C. hydrogeniphilus grows optimally at pH 5?5–6?5. In contrast to C. hydrogeniphilus, which is capable of poorheterotrophic growth on complex organic substrates, strainCRT is a strictly lithotrophic micro-organism. Thus, onthe basis of phylogenetic, morphological and physiological
features, it is proposed that CRT (=DSM 15016T=JCM 11957T) is the type strain of a novel species of Camini-bacter, for which the name Caminibacter profundus sp. nov.
Fig. 1. Electron micrographs of strain CRT. Negatively stained
cell showing polar flagellum (a) and ultrathin section of the cell(b). Bars, 0?5 mm.
The class ‘Epsilonproteobacteria’ (Garrity & Holt, 2001) isrepresented by a single tentative order, ‘Campylobacterales’.
The order presently contains the family Campylobacteraceae
(Vandamme & De Ley, 1991) and the as-yet tentative family
pH 6?9–7?0). Optimal NaCl concentration for growth was
‘Helicobacteraceae’ (Garrity & Holt, 2001). Levels of 16S
30 g l21; no growth was observed in media containing less
rRNA gene sequence similarity between the lineage encom-
than 5 or more than 50 g NaCl l21. Under optimal
passing Nautilia and Caminibacter and the ‘Campylo-
conditions, the doubling time was about 40 min and the cell
bacterales’ are about 83 % (Fig. 2). Phenotypic and genomic
yield reached 76108 cells ml21. A slightly higher cell yield
features also clearly distinguish the two phylogenetic
(about 1?56109 cells ml21) was obtained under 0?5 %
lineages (Table 1). It is therefore proposed that members
oxygen. Acetate, formate, butyrate, propionate, malate,
of the genera Nautilia and Caminibacter form a new order,
succinate, methanol, ethanol, pyruvate, lactate, fumarate,
Nautiliales ord. nov., represented by the single family
methylamine, glucose, sucrose, starch, peptone and yeast
extract did not support growth. Strain CRT did not growwhen sulfate, sulfite or thiosulfate were provided as alter-
Description of Nautiliales ord. nov.
native electron acceptors. To examine possible carbon
sources other than CO2, acetate, pyruvate, formate,methylamine, methanol and malate were tested; none of
Nautiliales (Nau.ti9li.a.les. N.L. fem. n. Nautilia the type
them supported growth. Strain CRT was sensitive to
genus of the order; N.L. -ales ending denoting an order; N.L.
rifampicin, vancomycin, penicillin and streptomycin (all at
fem. pl. n. Nautiliales the order of Nautilia).
Table 1. Differentiating characteristics of the families Nautiliaceae fam. nov., Campylobacteraceae and ‘Helicobacteraceae’
Data for Nautiliaceae were taken from Alain et al. (2002), Miroshnichenko et al. (2002) and this study. The family Campylobacteraceae con-tains the genera Campylobacter, Arcobacter, Sulfurospirillum and Thiovulum (data from Vandamme & De Ley, 1991; Vandamme et al., 1991;La Riviere & Schmidt, 1992; Schumacher et al., 1992; Luijten et al., 2003). The tentative family ‘Helicobacteraceae’ contains the generaHelicobacter and Wolinella (data from Tanner et al., 1981; Vandamme et al., 1991).
Helical, curved, S-shaped, spiral rods or ovoid
Microaerobic, some strains aerobic or anaerobic
Chemo-organotrophic; Sulfospirillum, mixotrophic;
*Sulfur is reduced by Sulfurospirillum species.
Order of the ‘Epsilonproteobacteria’ separate and distinct
sulfur or nitrate are used as electron acceptors. Chemo-
from the ‘Campylobacterales’. Segregation of these organ-
lithoautotrophs; mixotrophy occurs. Positive for H2 oxida-
isms into a new order is justified by (i) their distinct
tion. DNA G+C content of 29–35 mol%. Type genus:
phylogenetic position and (ii) their thermophilic way of
Nautilia Miroshnichenko et al. 2002.
life. Marine thermophilic rod-shaped bacteria, mean cellsize of 0?561?3 mm, non-spore-forming. Gram-negative.
Description of Nautiliaceae fam. nov.
Obligately anaerobic or microaerobic. For anaerobic growth,
Nautiliaceae (Nau.ti9li.a.ce.ae. N.L. fem. n. Nautilia the typegenus of the family; N.L. -aceae ending denoting a family;N.L. fem. pl. n. Nautiliaceae the family of Nautilia).
Description is the same as that for the order. Type genus:Nautilia Miroshnichenko et al. 2002.
Description of Caminibacter profundus sp. nov.
Caminibacter profundus (pro.fun9dus. L. masc. adj. profun-dus of the depths of the ocean).
Cells are motile, rod-shaped (1?2–1?560?5 mm) with singlepolar flagellum. Gram-negative cell wall structure. Anaero-bic to microaerobic. Spores absent. Moderate thermophile,growing at 45–65 uC (optimum 55 uC). Neutrophile, grow-ing at pH 6?5–7?4 (optimum pH 6?9–7?1). Grows in 5–50 gNaCl l21 (optimum around 30 g NaCl l21). Utilizes H2 asenergy source, elemental sulfur, nitrate or oxygen as electronacceptors and CO2 as carbon source. Nitrate and sulfur arerespectively reduced to ammonium and hydrogen sulfide inthe course of growth. Growth is not supported by acetate,
Fig. 2. Neighbour-joining dendrogram based on 16S rDNA
formate, butyrate, propionate, malate, succinate, methanol,
sequences showing the position of strain CRT in relation to its
ethanol, pyruvate, lactate, fumarate, methylamine, glucose,
phylogenetic neighbours, members of the genera Caminibacter
sucrose, starch, peptone or yeast extract. Acetate, pyruvate,
and Nautilia, ‘Epsilonproteobacteria’ and as-yet uncultured bac-teria from vents of the Pacific and Atlantic. Percentages of 500
formate, methylamine, methanol and malate cannot replace
bootstrap resamplings that support branching points above
CO2 as carbon source. Sulfate, sulfite and thiosulfate are not
70 % confidence are indicated. Bar, 10 nt substitutions per
utilized as electron acceptors. Grows in the presence of
100 sequence positions. The tree was rooted with 16S rDNA
chloramphenicol and tetracycline (both at 100 mg ml21).
sequences of members of the class ‘Gammaproteobacteria’.
DNA G+C content of the type strain is 32?1 mol%.
International Journal of Systematic and Evolutionary Microbiology 54
The type strain, CRT (=DSM 15016T=JCM 11957T), was
Luijten, M. L. G. C., de Weert, J., Smidt, H., Boschker, H. T. S.,
isolated from the content of a vent cap deployed in the
de Vos, W. M., Schraa, G. & Stams, A. J. M. (2003). Description of
Sulfurospirillum halorespirans sp. nov., an anaerobic tetrachloro-ethene-respiring bacterium, and transfer of Dehalospirillum multi-vorans to the genus Sulfurospirillum as Sulfurospirillum multivorans
comb. nov. Int J Syst Evol Microbiol 53, 787–793.
Maidak, B. L., Cole, J. R., Parker, C. T., Jr & 11 other authors (1999).
We thank Yves Fouquet (chief scientist) for inviting us to participate in
A new version of the RDP (Ribosomal Database Project). Nucleic
the Iris cruise (2001) and the crews of R/V L’Atalante and the pilots of
the ROV Victor. This work was supported by a CNRS/Rhoˆne-Poulenc
Miroshnichenko, M. L., Gongadze, G. M., Lysenko, A. M. &
grant, a PRIR grant from the Conseil Re´gional de Bretagne, INTAS
Bonch-Osmolovskaya, E. A. (1994). Desulfurella multipotens sp.
grant no. 99-1250 and the ‘Biodiversity’ programme of the Russian
nov., a new sulfur-respiring thermophilic eubacterium from Raoul
Ministry of Industry, Science and Technology. M. L. M. was supported
Island (Kermadec archipelago). Arch Microbiol 161, 88–93.
by the Ministe`re de l’Education Nationale during her stay in France.
Miroshnichenko, M. L., Kostrikina, N. A., L’Haridon, S., Jeanthon, C.,Hippe, H., Stackebrandt, E. & Bonch-Osmolovskaya, E. A. (2002).
Nautilia lithotrophica gen. nov., sp. nov., a thermophilic sulfur-reducing e-proteobacterium isolated from a deep-sea hydrothermal
Alain, K., Querellou, J., Lesongeur, F., Pignet, P., Crassous, P.,
vent. Int J Syst Evol Microbiol 52, 1299–1304.
Miroshnichenko, M. L., Kostrikina, N. A., Chernyh, N. A., Pimenov,
Caminibacter hydrogeniphilus gen. nov., sp. nov., a novel thermo-
N. V., Tourova, T. P., Antipov, A. N., Spring, S., Stackebrandt, E. &
philic, hydrogen-oxidizing bacterium isolated from an East Pacific
Bonch-Osmolovskaya, E. A. (2003). Caldithrix abyssi gen. nov., sp.
Rise hydrothermal vent. Int J Syst Evol Microbiol 52, 1317–1323.
nov., a nitrate-reducing, thermophilic, anaerobic bacterium isolated
Balch, W. E., Fox, G. E., Magrum, L. J., Woese, C. R. & Wolfe, R. S.
from a Mid-Atlantic Ridge hydrothermal vent, represents a novel
(1979). Methanogens: re-evaluation of a unique biological group.
bacterial lineage. Int J Syst Evol Microbiol 53, 323–329.
Moyer, C. L., Dobb, F. C. & Karl, D. M. (1995). Phylogenetic diversity
Bonch-Osmolovskaya, E. A., Sokolova, T. G., Kostrikina, N. A. &
of the bacterial community from a microbial mat at an active,
Zavarzin, G. A. (1990). Desulfurella acetivorans gen. nov. and
hydrothermal vent system. Appl Environ Microbiol 61, 1555–1562.
sp. nov. – a new thermophilic sulfur-reducing eubacterium. Arch
Polz, M. F. & Cavanaugh, C. M. (1995). Dominance of one bacterial
phylotype at a Mid-Atlantic Ridge hydrothermal vent site. Proc Natl
Campbell, B. J., Jeanthon, C., Kostka, J. E., Luther, G. W., III & Cary,
S. C. (2001). Growth and phylogenetic properties of novel bacteriabelonging to the epsilon subdivision of the Proteobacteria enriched
Rainey, F. A., Ward-Rainey, N., Kroppenstedt, R. M. & Stackebrandt, E.
from Alvinella pompejana and deep-sea hydrothermal vents. Appl
(1996). The genus Nocardiopsis represents a phylogenetically coherent
taxon and a distinct actinomycete lineage: proposal of Nocardiopsaceaefam. nov. Int J Syst Bacteriol 46, 1088–1092.
Desbruye`res, D. (1997). Molecular identification and localization
Reysenbach, A.-L., Longnecker, K. & Kirshtein, J. (2000). Novel
of filamentous symbiotic bacteria associated with the hydrothermal
bacterial and archaeal lineages from an in situ growth chamber
vent annelid Alvinella pompejana. Appl Environ Microbiol 63,
deployed at a Mid-Atlantic Ridge hydrothermal vent. Appl Environ
Corre, E., Reysenbach, A.-L. & Prieur, D. (2001). e-Proteobacterial
Schumacher, W., Kroneck, P. M. H. & Pfennig, N. (1992).
diversity from a deep-sea hydrothermal vent on the Mid-Atlantic
Comparative systematic study on ‘Spirillum’ 5175, Campylobacter
Ridge. FEMS Microbiol Lett 205, 329–335.
and Wolinella species. Arch Microbiol 158, 287–293.
Felsenstein, J. (1988). Phylogenies from molecular sequences:
Takai, K., Inagaki, F., Nakagawa, S., Hirayama, H., Nunoura, T.,
inference and reliability. Annu Rev Genet 22, 521–565.
Sako, Y., Nealson, K. H. & Horikoshi, K. (2003). Isolation and phylo-genetic diversity of members of previously uncultivated e-Proteobacteria
Garrity, G. M. & Holt, J. G. (2001). The road map to the Manual.
in deep-sea hydrothermal fields. FEMS Microbiol Lett 218, 167–174.
In Bergey’s Manual of Systematic Bacteriology, 2nd edn, vol. 1,pp. 119–166. Edited by D. R. Boone & R. W. Castenholz. New York:
Tanner, A. C. R., Badger, S. M., Lai, C.-H., Listgarten, M. A., Visconti,
R. A. & Socransky, S. S. (1981). Wolinella gen. nov., Wolinellasuccinogenes (Vibrio succinogenes Wolin et al.) comb. nov., and
Haddad, A., Camacho, F., Durand, P. & Cary, S. C. (1995).
description of Bacteroides gracilis sp. nov., Wolinella recta sp. nov.,
Phylogenetic characterization of the epibiotic bacteria associated
Campylobacter concisus sp. nov., and Eikenella corrodens from
with the hydrothermal vent polychaete Alvinella pompejana. ApplEnviron Microbiol 61, 1679–1687.
humans with periodontal disease. Int J Syst Bacteriol 31, 432–445.
Tru¨per, H. G. & Schlegel, H. G. (1964). Sulfur metabolism in
Jukes, T. H. & Cantor, C. R. (1969). Evolution of protein molecules. In Mammalian Protein Metabolism, vol. 3, pp. 21–132. Edited by
Thiorhodaceae. I. Quantitative measurements on growing cells of
H. N. Munro. New York: Academic Press.
Chromatium okenii. J Microbiol Serol 30, 225–232.
Vandamme, P. & De Ley, J. (1991).
La Riviere, J. W. M. & Schmidt, K. (1992). Morphologically
conspicuous sulfur-oxidizing eubacteria. In The Prokaryotes, 2nd
Campylobacteraceae. Int J Syst Bacteriol 41, 451–455.
edn, pp. 3934–3947. Edited by A. Balows, H. G. Tru¨per, M.
Vandamme, P., Falsen, E., Rossau, R., Hoste, B., Segers, P., Tytgat,
Dworkin, W. Harder & K.-H. Schleifer. New York: Springer.
R. & De Ley, J. (1991). Revision of Campylobacter, Helicobacter,
Longnecker, K. & Reysenbach, A. (2001). Expansion of geographic
and Wolinella taxonomy: emendation of generic descriptions and
distribution of a novel lineage of e-Proteobacteria to a hydrothermal
proposal of Arcobacter gen. nov. Int J Syst Bacteriol 41, 88–103.
vent site on the southern East Pacific Rise. FEMS Microbiol Ecol 35,
Wolin, E. A., Wolin, M. J. & Wolfe, R. S. (1963). Formation of methane
by bacterial extracts. J Biol Chem 238, 2882–2888.
SUGÁRTERÁPIÁS ÉS ONKOLÓGIAI SZAKMAI KOLLÉGIUM TITKÁR: PROF. DR. KÁSLER DR.SZŰCS MIKLÓS ORSZÁGOS ONKOLÓGIAI INTÉZET BÁCS-KISKUN 1122 BUDAPEST, RÁTH GYÖRGY U. 7-9. ONKORADIOLÓGIAI KÖZPONT TEL.: 224 8686, 224 8600 6000 KECSKEMÉT, NYÍRI ÚT 38. 0FAX.: 224 8687, 224 8620 TEL.:06-76-516-762 06-76-481-840 Javaslat az onkológiai l
To All MCPASD Staff Members: We are pleased to offer you a 403(b) plan to help you save for your future retirement on a tax sheltered basis. This is a good time to consider supplementing your retirement savings by enrolling in the 403(b) plan. Enclosed is a paper titled “Important Benefit Information Regarding Your 403(b) Plan”, which provides some general information. We are enclosi
 100 mg ml21). It grew in the presence of chloramphenicoland tetracycline (both at 100 mg ml21). The G+C contentof the DNA of isolate CRT was 32?1 mol%.
100 mg ml21). It grew in the presence of chloramphenicoland tetracycline (both at 100 mg ml21). The G+C contentof the DNA of isolate CRT was 32?1 mol%.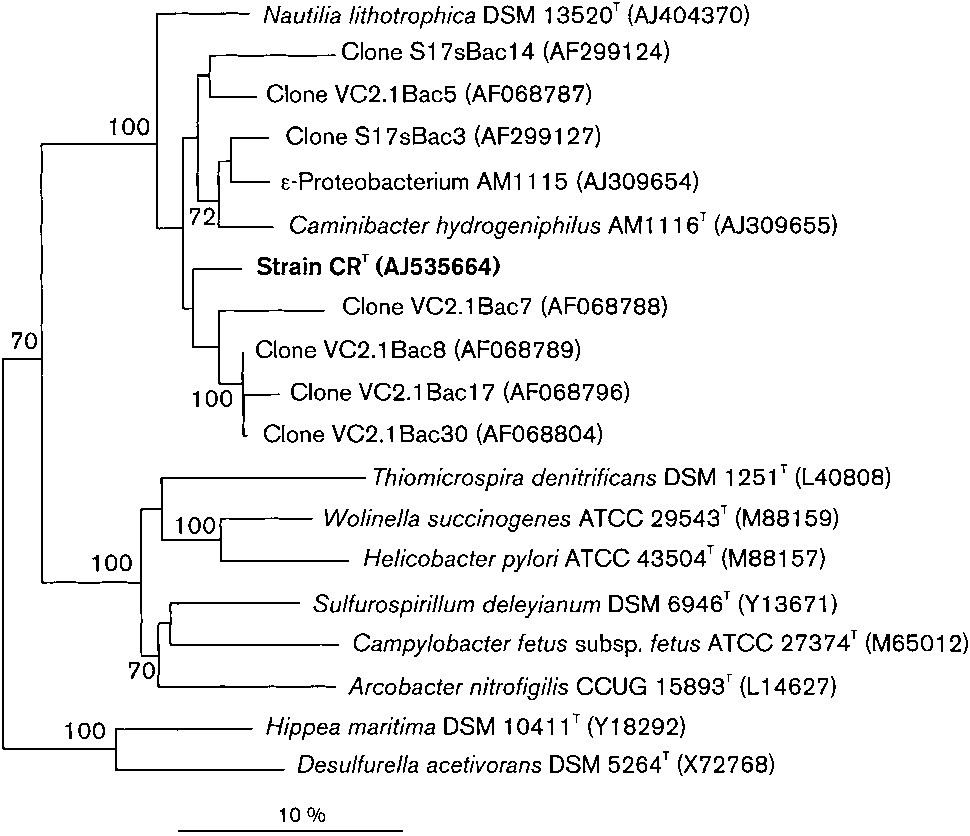 Table 1. Differentiating characteristics of the families Nautiliaceae fam. nov., Campylobacteraceae and ‘Helicobacteraceae’
Data for Nautiliaceae were taken from Alain et al. (2002), Miroshnichenko et al. (2002) and this study. The family Campylobacteraceae con-tains the genera Campylobacter, Arcobacter, Sulfurospirillum and Thiovulum (data from Vandamme & De Ley, 1991; Vandamme et al., 1991;La Riviere & Schmidt, 1992; Schumacher et al., 1992; Luijten et al., 2003). The tentative family ‘Helicobacteraceae’ contains the generaHelicobacter and Wolinella (data from Tanner et al., 1981; Vandamme et al., 1991).
Table 1. Differentiating characteristics of the families Nautiliaceae fam. nov., Campylobacteraceae and ‘Helicobacteraceae’
Data for Nautiliaceae were taken from Alain et al. (2002), Miroshnichenko et al. (2002) and this study. The family Campylobacteraceae con-tains the genera Campylobacter, Arcobacter, Sulfurospirillum and Thiovulum (data from Vandamme & De Ley, 1991; Vandamme et al., 1991;La Riviere & Schmidt, 1992; Schumacher et al., 1992; Luijten et al., 2003). The tentative family ‘Helicobacteraceae’ contains the generaHelicobacter and Wolinella (data from Tanner et al., 1981; Vandamme et al., 1991).