Tadalafil entfaltet seine Wirkung über eine selektive Hemmung der PDE5, wodurch die Konzentration von cGMP im glatten Muskelgewebe stabil bleibt. Diese biochemische Modulation resultiert in einer langanhaltenden Relaxation der Gefäßwände. Der Wirkstoff wird nach oraler Einnahme effizient resorbiert, mit einer Bioverfügbarkeit von rund 80 %. Seine Halbwertszeit von bis zu 36 Stunden ist innerhalb dieser Substanzklasse außergewöhnlich. Abgebaut wird er in der Leber, hauptsächlich durch CYP3A4, mit anschließender biliärer Exkretion. Typische unerwünschte Wirkungen entstehen durch eine verstärkte Vasodilatation, etwa Kopfschmerzen oder Flush. Pharmakologisch wird cialis generika vor allem durch die verlängerte Wirkungsdauer charakterisiert.
Secure.footprint.net
Gatorade Sports Science 61 Institute ® CARBOHYDRATES, BRANCHED-CHAIN AMINO ACIDS AND ENDURANCE: THE CENTRAL FATIGUE HYPOTHESIS
This article is a modification of the original article that appeared in a supplemental issue to the International Journal of SportsNutrition, Vol. 5, 1995. The supplement represented the proceedings of the Gatorade Sports Science Institute Ergogenic AidsConference, held in Chicago, November, 1994. J. Mark Davis Department of Exercise Science The University of South Carolina Columbia, SC 29208 KEY POINTS 1. The mechanisms of central fatigue are largely unexplored, but the central fatigue hypothesis suggests that increased brain sero-
tonin (5-HT) can cause a deterioration in sport and exercise performance. There is now convincing evidence thatexercise-induced increases in the plasma free tryptophan (f-TRP)/branched-chain amino acids (BCCA) ratio are associated withincreased brain 5-HT and the onset of fatigue during prolonged exercise.
2. When drugs are administered to alter brain 5-HT, they have the predicted effects on exercise performance. The influence of
nutritional manipulations of f-TRP/BCCA on performance is less well established.
3. The effects of BCCA supplementation on exercise performance are mixed, and the published studies often suffer from method-
ological flaws. Alternatively, dramatic reductions in f-TRP/BCCA and enhanced performance accompany carbohydrate feedingsduring prolonged exercise. However, it is difficult to distinguish between the effects of carbohydrate feedings on mechanismsthat reside in the brain versus the muscles themselves. INTRODUCTION
Muscular fatigue is commonly defined as a failure to maintain the required or expected force or power output (22). The causes
of muscular fatigue involve specific impairments within the muscle itself, including transmission of the neural stimulus to themuscle at the motor end plate and propagation of that stimulus throughout the muscle (35, 38), disruption of calcium release anduptake within the sarcoplasmic reticulum (23), substrate depletion (19), and various other metabolic events that impair energyprovision and muscle contraction (27). Fatigue can also result from alterations within the central nervous system (CNS) althoughessentially nothing is known about the specific mechanisms underlying this type of fatigue (central fatigue).
The potential role of central fatigue during prolonged exercise has received very little scientific attention, even though it is well
known that “psychological factors” can affect exercise performance (3). In fact, the lack of adequate CNS drive to the workingmuscles is the most likely explanation of fatigue in most people during normal activities. Furthermore, the often debilitating fatiguethat accompanies viral or bacterial infections, recovery from injury or surgery, chronic fatigue syndrome, and depression almostcertainly cannot be explained by a dysfunction within the muscles themselves. Fatigue under these circumstances probablyinvolves the CNS, but the specific causes have not been elucidated.
An important role for central fatigue during exercise in healthy people has been suggested in a number of studies (3, 9, 34),
but most of these investigations have failed to provide plausible physiological mechanisms. Recently, however, interestingnew theories have been proposed that implicate various neurotransmitters such as serotonin (5-hydroxytryptamine, or 5-HT)(32), norepinephrine, and dopamine (8, 28, 30, 33) in central fatigue during exercise. This review will focus primarily on thescientific evidence regarding brain 5-HT as a potential mediator of central fatigue during prolonged exercise. The exciting, butmore limited, evidence that proper nutrition may be able to alter brain 5-HT synthesis and delay central fatigue will also bediscussed. For this review, a working definition of central fatigue will be used which suggests that central fatigue is a subsetof fatigue that is associated with specific alterations in CNS function and that cannot reasonably be explained by peripheralmarkers of muscle fatigue. BRAIN 5-HT AND THE CENTRAL FATIGUE HYPOTHESIS
(32). There is a large body of literature
increase when there is an increase in the
primary factor leading to this response.
fatty acids (FFAs) increase in the blood,
cise and thereby cause a deterioration in
(˜180 min. run time) and untrained (˜72
sport and exercise performance (32).
increases in brain concentrations of 5-HT
after 1 hr. of treadmill exercise, and at
VO2max in rats (20 m • min-1, 5% grade).
After 1 hr. of exercise, the concentrations
even further in the midbrain andstriatum. DA and DOPAC also increasedin the midbrain, striatum, and hypothal-
ASSOCIATION BETWEEN FATIGUE AND INCREASES IN
to baseline levels at fatigue. These results
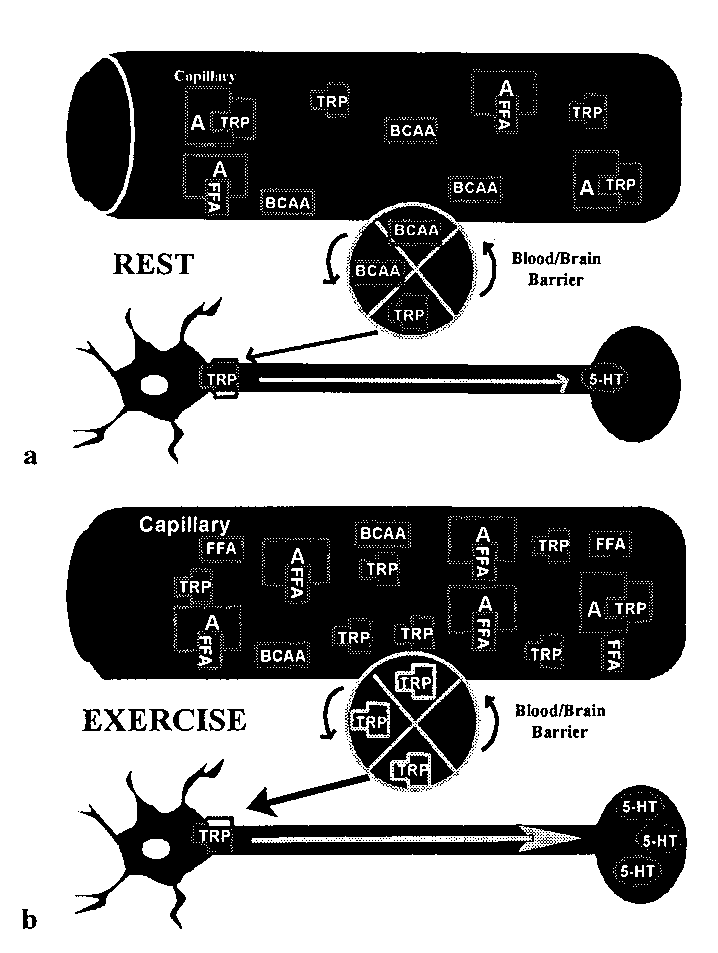
Figure 1: Primary components of the central fatigue BRAIN 5-HT AND 5-HIAA
hypothesis at rest and during prolonged exercise. (a)
The situation at rest with regard to plasma concentra-
tions of branched-chain amino acids (BCAAs), free
appears to peak at the time of fatigue.
fatty acids (FFAs), and tryptophan (TRP) (bound and
Chaouloff (14-18) and Newsholme (13).
unbound to albumin) and their proposed effects on
transport of TRP across the blood-brain barrier for the
that is usually associated with increased
synthesis of serotonin (5-HT) in serotonergic neurons. (b) What happens during prolonged exercise to
increase the synthesis of 5-HT in the brain and lead to
central fatigue. Adapted from J.D. Fernstrom. Dietary
prolonged exercise as fatigue develops.
amino acids and brain function. J. Am. Diet. Assoc.
The significance of this apparent inverse
EXPERIMENTAL ALTERATIONS OF BRAIN 5-HT ACTIVITY
that this may cause central fatigue.
central fatigue may occur as a result.
(5-HT antagonists) would delay fatigue.
f-TRP to the brain for 5-HT synthesis.
hypothesis is that a low ratio of 5-HT:DA
athletes’ actual situations, such studies
are often limited in scientific value. For
NUTRITIONAL EFFECTS ON 5-HT AND CENTRAL FATIGUE
or other factors. In addition, studies of
treatments to prevent bias on the part of
these studies often fail to control impor-
tant variables such as exercise intensity
time to fatigue occurred earlier (41) and
effects, and there were no differences in
slightly lower (˜19%) after the race.
between the drug and placebo trials. SUMMARY AND CONCLUSIONS
exercise test to fatigue. Verger et al.
mechanisms have not been established.
their peaks at fatigue, (b) the increase in
severe symptoms of central fatigue (7).
peripheral markers of muscle fatigue.
subjects drank 5 ml • kg-1 • hr-1 of either
a role in delaying central fatigue during
prolonged exercise, the scientific data in
tion to plasma free fatty acids), whereas
are reasons to believe that this approach
strategy. In this case, however, it is not
tions and to decrease the likelihood that
large attenuation (fivefold to sevenfold)
edge in this area is rudimentary at best.
moregulatory, and metabolic functions. References
1. Akil, H., and J.C. Liebeskind. Monoaminergic mechanisms of stimulation-produced analgesia. Brain Res. 94:279-296, 1975.
2. Alper, R.H. Evidence for central and peripheral serotonergic control of corticosterone secretion in the conscious rat. Neuroendocrinology 51:255-260, 1990.
3. Asmussen, E. Muscle fatigue. Med. Sci. Sports. 11(4): 313-321, 1979.
4. Bailey, S.P., J.M. Davis, and E.N. Ahlborn. Effect of increased brain serotonergic (5-HT1C) activity on endurance performance in the rat. Acta Physiol. Scand. 145 (91): 75-76, 1992.
5. Bailey, S.P., J.M., Davis, and E.N. Ahlborn. Brain serotonergic activity affects endurance performance in the rat. Int. J. Sports Med. 6:330-333, 1993.
6. Bailey, S.P., J.M. Davis, and E.N. Ahlborn. Neuroendocrine and substrate responses to altered brain 5-HT activity during prolonged exercise to fatigue. J. Appl. Physiol.
7. Banister, W.W., and B.J.C. Cameron. Exercise-induced hyperammonemia: Peripheral and central effects. Int. J. Sports Med. 11 (Suppl. 2):S129-S142, 1990.
8. Bhagat, B., and N. Wheeler. Effect of amphetamine on the swimming endurance of rats. Neuropharmacology, 12:711-713, 1973.
9. Bigland-Ritchie, B., F. Furbush, and J.J. Woods. Fatigue of intermittent submaximal voluntary contractions: Central and peripheral factors. J. Appl. Physiol. 61 (2):421-429, 1986.
10. Blomstrand, E., F. Celsing, and E.A. Newsholme. Changes in plasma concentrations of aromatic and branch-chain amino acids during sustained exercise in man and their possible
role in fatigue. Acta Physiol. Scand. 133:115-121, 1988.
11. Blomstrand, E., P. Hassmen, B. Ekblom, and E.A. Newsholme. Administration of branched-chain amino acids during sustained exercise—Effects on performance and on plasma
concentration of some amino acids. Eur. J. Appl. Physiol. 63:83-88, 1991.
12. Blomstrand, E., P. Hassmen, and E.A. Newsholme. Effect of branched-chain amino acid supplementation on mental performance. Acta Physiol. Scand. 136:473-481, 1991.
13. Blomstrand, E., D. Perrett, M. Parry-Billings, and E.A. Newsholme. Effect of sustained exercise on plasma amino acid concentrations and on 5-hydroxytryptamine metabolism in
six different brain regions in the rat. Acta Physiol. Scand. 136:473-481, 1989.
14. Chaouloff, F., J.L. Elghozi, Y. Guezennec, and D. Laude. Effects of conditioned running on plasma, liver and brain tryptophan and on brain 5-hydroxytryptamine metabolism of the
rat. Brit. J. Pharm. 86:33-41, 1985.
15. Chaouloff, F., G.A. Kennett, B. Serrurier, D. Merina, and G. Curson. Amino acid analysis demonstrates that increased plasma free tryptophan causes the increase of brain trypto-
phan during exercise in the rat. J. Neurochem. 46:1647-1650, 1986.
16. Chaouloff, F., D. Laude, and J.L. Elghozi. Physical exercise: Evidence for differential consequences of tryptophan on 5-HT synthesis and metabolism in central serotonergic cell
bodies and terminals. J. Neural Trans. 78:121-130, 1989.
17. Chaouloff, F., D. Laude, Y. Guezennec, and J.L. Elghozi. Motor activity increases tryptophan, 5-hydroxyindoleacetic acid, and homovanillic acid in ventricular cerebrospinal fluid
of the conscious rat. J. Neurochem. 46:1313-1316, 1986.
18. Chaouloff, F., D. Laude, D. Merino, B. Serrurier, Y. Guezennec, and J.L. Elghozi. Amphetamine and alpha-methyl-p-tyrosine affect the exercise induced imbalance between the
availability of tryptophan and synthesis of serotonin in the brain of the rat. Neuropharmacology 26:1099-1106, 1987.
19. Coggan, A.R., and E.F. Coyle. Carbohydrate ingestion during prolonged exercise: Effects on metabolism and performance. Exer. Sport Sci. Rev. 19:1-40, 1991.
20. Davis, J.M., S.P. Bailey, D.A. Jackson, A.B. Strasner, and SlL. Morehouse. Effects of a serotonin (5-HT) agonist during prolonged exercise to fatigue in humans. Med. Sci. Sports
21. Davis, J.M., S.P. Bailey, J.A. Woods, F.J. Galiano, M. Hamilton, and W.P. Bartoli. Effects of carbohydrate feedings on plasma free-tryptophan and branched-chain amino acids
during prolonged cycling. Eur. J. Appl. Physiol. 65:513-519, 1992.
22. Enoka, R.M., and D.G. Stuart. Neurobiology of muscle fatigue. J. Appl. Physiol. 72(5):1631-1648, 1992.
23. Fitts, R.H., and J.M. Metzger. Mechanisms of muscular fatigue. In Principles of Exercise Biochemistry (2nd ed., Vol. 38), J.R. Poortmans (Ed.), Basel, Switzerland: Karger, 1993,
24. Freed, C.R., and B.K. Yamamoto. Regional brain dopamine metabolism: A marker for speed, direction, and posture of moving animals. Science 229:62-65, 1985.
25. Galiano, F.J., J.M. Davis, S.P. Bailey, J.A. Woods, and M. Hamilton. Physiologic, endocrine and performance effects of adding branch chain amino acids to a 6% carbohydrate-
electrolyte beverage during prolonged cycling. Med. Sci. Sports Exerc. 23(4):S14, 1991.
26. Gerald, M.C. Effect of (+)- amphetamine on the treadmill endurance performance of rats. Neuropharmacology 17:703-704, 1978.
27. Green, H.J. Neuromuscular aspects of fatigue. Can. J. Sports Sci. 12 (Suppl. 1):7s-19s, 1987.
28. Heyes, M.P., E.S. Garnett, and G. Coates. Central dopaminergic activity influences rats’ ability to exercise. Life Sci. 36:671-677, 1985.
29. Jauvet, M. and J-F. Pujol. Effects of central alterations of serotonergic neurons upon the sleep-waking cycle. Adv. Biochem. Psychopharm. 11:199-209, 1974.
30. Laties, V.G., and B. Weiss. The amphetamine margin in sports. Federation Proc. 40:2689-2692, 1981.
31. Leathwood, P.D., and J.D. Fernstrom. Effect of an oral tryptophan/carbohydrate load on tryptophan, large neutral amino acid, and serotonin and 5-hydroxyindoleacetic acid levels in
monkey brain. J. Neural Tran. 79:25-34, 1990.
32. Newsholme, E.A., I.N. Acworth, and E. Blomstrand. Amino acids, brain neurotransmitters and a functional link between muscle and brain that is important in sustained exercise. In
Advances in Myochemistry, G. Benzi (Ed.), London: John Libbey Eurotext, 1987, pp. 127-133.
33. Salter, C.A. Dietary tyrosine as an aid to stress resistance among troops. Military Med. 154:144-146, 1989.
34. Secher, N.H. Central nervous influence on fatigue. In Endurance in Sport, R.J. Shephard and P.-O. Astrand (Eds.), Boston: Blackwell Scientific, 1992, pp. 96-106.
35. Sjogaard, G. Muscle fatigue. Med Sports Sci. 26:98-109, 1987.
36. Varnier, M., P. Sarto, D. Martines, L. Lora, F. Carmignoto, G. Leese, and R. Naccarato. Effect of infusing branched-chain amino acid during incremental exercise with reduced
muscle glycogen content. Eur. J. Appl. Physiol. 69:26-31, 1994.
37. Verger, P.H., P. Aymard, L. Cynobert, G. Anton, and R. Luigi. Effects of administration of branched-chain amino acids vs. glucose during acute exercise in the rat. Physiol. Behav.
38. Vollestad, N.K., and O.M. Sejersted. Biochemical correlates of fatigue. Eur. J. Appl. Physiol. 57:336-347, 1988.
39. Wagenmakers, A.J.M., E.J. Bechers, F. Brouns, H. Kuipers, P.B. Soeters, G.J. Van der Vusse, and W.H.M. Saris. Carbohydrate supplementation, glycogen depletion, and amino
acid metabolism during exercise. Am. J. Physiol. 260:E883-E890, 1991.
40. Wagenmakers, A.J.M., J.H. Coakley, and R.H.T. Edwards. Metabolism of branched-chain amino acids and ammonia during exercise: Clues from McArdle’s disease. Int. J. Sports
41. Wilson , W.M., and R.J. Maughan. Evidence for a possible role of 5-hydroxytryptamine in the genesis of fatigue in man: Administration of paroxetine, a 5-HT re-uptake inhibitor,
reduces the capacity to perform prolonged exercise. Exp. Physiol. 77:921-924, 1992.
42. Young, S.N. The clinical psychopharmacology of tryptophan. In Nutrition and the Brain (Vol. 7), R.J. Wurtman and J.J. Wurtman (Eds.), New York: Raven, 1986, pp. 49-88.
Pressemitteilung Nr. 2/2008 - Nürnberg, 07. April 2008 STUDIO GONG und SOMA 2 vermarkten gemeinsam Um das Onlineangebot der STUDIO GONG GmbH & Co. Studiobetriebs KG zu erweitern, arbeitet der führende Hörfunkvermarkter mit dem Onlinevermarkter SOMA 2 GmbH zusammen. Die Kooperation stärkt die Position der beiden Unternehmen im Bereich Radio und Regionalwerbung sowie der nationalen
 BRAIN 5-HT AND THE CENTRAL
BRAIN 5-HT AND THE CENTRAL