Tadalafil entfaltet seine Wirkung über eine selektive Hemmung der PDE5, wodurch die Konzentration von cGMP im glatten Muskelgewebe stabil bleibt. Diese biochemische Modulation resultiert in einer langanhaltenden Relaxation der Gefäßwände. Der Wirkstoff wird nach oraler Einnahme effizient resorbiert, mit einer Bioverfügbarkeit von rund 80 %. Seine Halbwertszeit von bis zu 36 Stunden ist innerhalb dieser Substanzklasse außergewöhnlich. Abgebaut wird er in der Leber, hauptsächlich durch CYP3A4, mit anschließender biliärer Exkretion. Typische unerwünschte Wirkungen entstehen durch eine verstärkte Vasodilatation, etwa Kopfschmerzen oder Flush. Pharmakologisch wird cialis generika vor allem durch die verlängerte Wirkungsdauer charakterisiert.
Untitled
International Journal of Systematic and Evolutionary Microbiology (2006), 56, 2927–2930
Cyclobacterium lianum sp. nov., a marinebacterium isolated from sediment of an oilfield inthe South China Sea, and emended description ofthe genus Cyclobacterium
Jiao-Yan Ying,1,2 Bao-Jun Wang,2 Su-Sheng Yang1and Shuang-Jiang Liu2
College of Biological Sciences, China Agricultural University, Beijing 100094, People’s
2State Key Laboratory of Microbial Resources, Institute of Microbiology, Chinese Academy of
Sciences, ZhongGuanCun, Haidian, Beijing 100080, People’s Republic of China
The marine bacterial strain HY9T was isolated from sediment from the South China Sea. StrainHY9T is aerobic, heterotrophic and rose-pigmented. The cells are non-motile and curved, i.e. ring-like or horseshoe-shaped. The 16S rRNA gene sequence of strain HY9T was determined and
BLAST searches revealed that it possessed significant sequence similarities with respect to
Cyclobacterium species (92?8–93?6 %). Phylogenetic analysis confirmed that strain HY9T wastightly clustered with members of the genus Cyclobacterium. The cellular morphology andchemotaxonomic and phenotypic properties of strain HY9T showed that it should be classified as amember of the genus Cyclobacterium. Significant evolutionary distances and a range of phenotypicfeatures distinguished strain HY9T from previously described Cyclobacterium species. Hence,strain HY9T represents a novel species in the genus Cyclobacterium, for which the nameCyclobacterium lianum sp. nov. is proposed. The type strain is HY9T (=CGMCC 1.6102T=JCM14011T). On the basis of this study and previously described properties of Cyclobacteriumspecies, an emended description of the genus Cyclobacterium is proposed.
Many members of the phylum Bacteroidetes (or Cytophaga–
designated strain HY9T and isolated from sediment of the
Flavobacterium–Bacteroides) are involved in biogeochemical
South China Sea. A polyphasic analysis of strain HY9T
cycles in aquatic environments (Bowman et al., 2003; and
indicated that the isolate represents a novel species of the
references therein). For instance, members of the genus
Cyclobacterium displaying the unique ring-like and horse-shoe-shaped morphology are common constituents of
Strain HY9T was isolated from sediment of the Xijiang
marine environments. At the time of writing, the genus
oilfield in the South China Sea, near Fujian Province, China;
Cyclobacterium (Raj & Maloy, 1990) contains two species
sampling was performed at a depth of about 100 m. For
with validly published names: Cyclobacterium marinum,
isolation, serially diluted sediment samples were spread onto
isolated from the deep sea in the Gulf of California (Raj
low-organic marine agar 2216 plates [containing 0?5 g
& Maloy, 1990; Euze´by, 1998), and Cyclobacterium
peptone l21 and 0?1 g yeast extract l21; the salt composition
amurskyense, isolated from water in the Sea of Japan
and concentration were the same as in marine agar 2216
(Nedashkovskaya et al., 2005). Organisms sharing high
(MA; Difco)] and incubated at 30 uC for 10 days. A colony
levels of 16S rRNA gene sequence similarity with Cyclo-
of HY9T was collected and subcultured on MA. The
bacterium species have also been found in salt-marsh
temperature and pH ranges for growth, the tolerance of
sediment (GenBank accession no. AY259502) and a soda
NaCl and the substrates used as sole carbon sources were
lake (AF275712). In this study, we describe a rose-pigmented
determined according to the methods described by Cho &
bacterial strain with ring-like and horseshoe-shaped cells,
Giovannoni (2003). Strain HY9T was tested on API ZYM(bioMe´rieux) and Biolog GN2 MicroPlate systems, accord-ing to the manufacturers’ instructions. All other biochem-
The GenBank/EMBL/DDBJ accession number for the 16S rRNA genesequence of strain HY9T is DQ534063.
ical tests were performed according to Dong & Cai (2001). Susceptibility to antibiotics was determined using filter-
A transmission electron micrograph of strain HY9T, two phylogenetictrees and a table of fatty acid compositions are available as
paper discs containing various antibiotics, as specified in the
supplementary material in IJSEM Online.
species description. The morphology of cells grown on MA
Table 1. Differential properties of strain HY9T and the type
27F and 1492R (Lane, 1991) and sequenced. Preliminary
comparisons with sequences held in GenBank, performedusing BLASTN searches, revealed that the isolate was related
Strains: 1, HY9T; 2, C. marinum LMG 13164T; 3, C. amurskyense
to C. marinum. Further analysis of 16S rRNA gene sequences
KMM 6143T. Data are from Nedashkovskaya et al. (2005) and
was performed using MEGA, version 3.1 (Kumar et al., 2004).
Strain HY9T showed highest sequence similarity with C.
marinum DSM 745T (93?6 %) and C. amurskyense KMM6143T (92?8 %), followed by members of the genera
(87?8–90?3 %), Chimaereicella (88?4 %) and Algoriphagus
(88?3–89?5 %). Phylogenetic trees were constructed with
MEGA, version 3.1, using the neighbour-joining method
(Saitou & Nei, 1987), maximum parsimony (Fitch, 1971)
and minimum evolution (Felsenstein, 1997). The resulting
tree topology was evaluated using the Kimura two-
parameter calculation model (Kumar et al., 2004) based
on 1000 replicates. As shown in Fig. 1 and Supplementary
Fig. S2 (available in IJSEM Online), strain HY9T formed a
monophyletic clade with C. marinum and C. amurskyense.
The closest relatives of this clade were members of the genera
Aquiflexum and Belliella (Brettar et al., 2004a, b).
The whole-cell fatty acid composition was determined from
cells cultured at 30 uC on MA for 2 days. The cellular fatty
acids were extracted, methylated and analysed by using the
Sherlock Microbial Identification System (MIDI) according
to the manufacturer’s instructions. The major cellular fattyacids were iso-C15 : 0 (28?3 %), summed feature 3 (iso-C15 : 02-OH and/or C
for 2 days at 30 uC was studied using transmission electron
16 : 1v7c; 16?6 %), iso-C17 : 1v9c (10?3 %),
17 : 0 3-OH (8?0 %) and anteiso-C15 : 0 (6?4 %), similar
to the profiles reported for C. marinum and C. amurskyense
The novel isolate displayed the basic characteristics of
(Nedashkovskaya et al., 2005). The complete fatty acid
members of the genus Cyclobacterium, e.g. the colonies were
composition of strain HY9T is given in Supplementary
rose-pigmented and the cells were curved, ring-like or
Table S1 (available in IJSEM Online). The G+C content of
horseshoe-shaped (see Supplementary Fig. S1 available in
the DNA was determined by thermal denaturation (Marmur
IJSEM Online). Other phenotypic properties of strain HY9T
& Doty, 1962; Seidler & Mandel, 1971) using DNA from
are given in the species description and in Table 1.
Escherichia coli K-12 as a control. The G+C content ofstrain HY9T was 45?2 mol%, which is slightly higher than
The almost-complete 16S rRNA gene (1485 bp) of strain
those of the type strains of C. marinum (41?9 mol%) and C.
HY9T was amplified using the bacterial universal primers
amurskyense (41?3 mol%) (Nedashkovskaya et al., 2005).
sequences of strain HY9T and related mem-bers
Bootstrap values (expressed as percentagesof 1000 replicates) greater than 50 % areshown at branching points. Bar, 0?02 substi-tutions per nucleotide position.
International Journal of Systematic and Evolutionary Microbiology 56
On the basis of these results, we concluded that strain HY9T
1-phosphate are oxidized. Weak or variable results are
represents a novel species within the genus Cyclobacterium,
detected with glycogen, N-acetyl-D-galactosamine, i-ery-
for which the name Cyclobacterium lianum sp. nov. is
thritol, D-mannitol, D-psicose, L-rhamnose, D-sorbitol,
proposed. The properties that serve to differentiate strain
monomethyl succinate, D-gluconic acid, a-ketovaleric
HY9T, C. marinum and C. amurskyense from each other are
acid, succinic acid, L-alanyl glycine, L-asparagine, L-aspartic
listed in Table 1. As some of the properties of C. lianum
acid, glutamic acid, L-ornithine, L-proline, L-pyroglutamic
(from this study) and C. amurskyense (Nedashkovskaya
acid, DL-serine, L-threonine, c-aminobutyric acid, uridine,
et al., 2005) were not included in or are not compatible with
2-aminoethanol and glucose 6-phosphate. Resistant to the
the original description of the genus Cyclobacterium (Raj &
following antibiotics (mg): gentamicin (10), neomycin (30),
Maloy, 1990), an emended description of the genus is also
polymyxin B (300), streptomycin (10) and tetracycline (30).
Sensitive to the following antibiotics (mg): ampicillin (10),carbenicillin (100), vancomycin (30), ciprofloxacin (5),
Description of Cyclobacterium lianum sp. nov.
rifampicin (5), norfloxacin (10), chloramphenicol (30),benzyl penicillin (10), kanamycin (30) and erythromycin
Cyclobacterium lianum (N.L. neut. adj. lianum pertaining to
(15). The major cellular fatty acids (>5 %) are iso-C
Li, named in honour of Professor Ji-Lun Li, who devotes
himself to microbiological research and education in
16 : 1v7c; 16?6 %), iso-C17 : 1v9c (10?3 %), iso-C17 : 0 3-OH
(8?0 %) and anteiso-C15 : 0 (6?4 %). The molar G+C content
Cells are Gram-negative, aerobic and heterotrophic, non-
motile, curved, ring-like or horseshoe-shaped, 0?4–0?5 mm
The type strain, HY9T (=CGMCC 1.6102T=JCM 14011T),
wide, and the outer diameter of rings is 1?5–1?8 mm.
was isolated from sediment from the Xijiang oilfield in the
Colonies grown for 3 days on MA are circular (2–3 mm in
diameter), light rose in colour and shiny. Growth occurs at15–42 uC (optimum 33 uC), at pH 6?5–9?0 (optimum
pH 7?5–8?0) and with 0?1–12 % NaCl (optimum 1–4 %).
Positive for oxidase and catalase activities, but negative forarginine dihydrolase, urease and lecithinase activities.
Colonies on MA are pink-pigmented and shiny. Cells are
Indole and H2S are not produced and nitrate is not reduced.
curved, ring-like or horseshoe-shaped. Neutrophilic and
Aesculin and Tween 20 are hydrolysed. Tweens 40 and 80 are
mesophilic. Optimal growth temperature range is 25–30 uC.
hydrolysed weakly; agar, casein, gelatin, starch, DNA and
NaCl is required for growth. The major cellular fatty acids
carboxymethyl-cellulose are not hydrolysed. Glucose,
are iso-C15 : 0, summed feature 3 (iso-C15 : 0 2-OH and/or
sucrose, D-melibiose, ribose, lactose, galactose, maltose,
C16 : 1v7c), iso-C17 : 1v9c, iso-C17 : 0 3-OH and anteiso-C15 : 0.
melezitose, inulin, L-rhamnose, L-arabinose, D-raffinose,
The DNA G+C content is 41–45 mol%.
trehalose, cellobiose, methyl a-D-glucoside and gluconateare utilized as sole carbon sources. Glycerol, mannitol, D-mannose, D-fructose, D-xylose, lactic acid, succinate,
malate, pyruvate and L-glutamic acid are weakly utilized.
This work was supported by grants from the National Natural Science
L-Fucose, L-sorbose, dulcitol, adonitol, myo-inositol, citrate,
malonate, L-lysine, L-alanine, formic acid, butyric acid andcaprate are not utilized. Forms acid from glucose, ribose(weakly), sucrose, D-melibiose, lactose, galactose, maltose,
melezitose, inulin, L-rhamnose, L-arabinose, D-raffinose,
Bowman, J. P., Nichols, C. M. & Gibson, J. A. E. (2003). Algoriphagus
trehalose, cellobiose, D-xylose, glycerol (weakly) and methyl
ratkowskyi gen. nov., sp. nov., Brumimicrobium glaciale gen. nov., sp.
a-D-glucoside. Shows strong activity in the API ZYM
nov., Cryomorpha ignava gen. nov., sp. nov. and Crocinitomix
system for alkaline and acid phosphatases, leucine and
catalasitica gen. nov., sp. nov., novel flavobacteria isolated from
valine arylamidases, naphthol-AS-BI-phosphohydrolase, b-
various polar habitats. Int J Syst Evol Microbiol 53, 1343–1355.
galactosidase, a- and b-glucosidases and N-acetyl-b-gluco-
Brettar, I., Christen, R. & Ho¨fle, M. G. (2004a). Belliella baltica gen.
saminidase. Shows weak activity for esterases C4 and C8,
nov., sp. nov., a novel marine bacterium of the Cytophaga–
Flavobacterium–Bacteroides group isolated from surface water of
the central Baltic Sea. Int J Syst Evol Microbiol 54, 65–70.
activity is shown for trypsin, a-chymotrypsin, b-glucur-
Brettar, I., Christen, R. & Ho¨fle, M. G. (2004b). Aquiflexum balticum
onidase, a-fucosidase or lipase (C14). In GN2 MicroPlates,
gen. nov., sp. nov., a novel marine bacterium of the Cytophaga–
dextrin, N-acetyl-D-glucosamine, L-arabinose, D-cellobiose,
Flavobacterium–Bacteroides group isolated from surface water of the
D-fructose, D-galactose, gentiobiose, a-D-glucose, a-D-
central Baltic Sea. Int J Syst Evol Microbiol 54, 2335–2341.
Cho, J. C. & Giovannoni, S. J. (2003). Parvularcula bermudensis gen.
methyl b-D-glucoside, D-raffinose, sucrose, D-trehalose,
nov., sp. nov., a marine bacterium that forms a deep branch in the a-
turanose, D-galacturonic acid, DL-lactic acid, glucurona-
Proteobacteria. Int J Syst Evol Microbiol 53, 1031–1036.
mide, L-alaninamide, L-alanine, DL-carnitine, 2,3-butane-
Dong, X.-Z. & Cai, M.-Y. (2001). Determinative Manual for Routine
Bacteriology. Beijing: Scientific Press.
Euze´by, J. P. (1998). Taxonomic note: necessary correction of
Nedashkovskaya, O. L., Kim, S. B., Lee, M. S., Park, M. S., Lee, K. H.,
specific and subspecific epithets according to Rules 12c and 13b of
Lysenko, A. M., Oh, H. W., Mikhailov, V. V. & Bae, K. S. (2005).
the International Code of Nomenclature of Bacteria (1990 Revision).
Cyclobacterium amurskyense sp. nov., a novel marine bacterium
Int J Syst Bacteriol 48, 1073–1075.
isolated from sea water. Int J Syst Evol Microbiol 55, 2391–2394.
Felsenstein, J. (1997). An alternative least-squares approach to
Raj, H. D. & Maloy, S. R. (1990). Proposal of Cyclobacterium
inferring phylogenies from pairwise distances. Syst Biol 46, 101–111.
marinum gen. nov., comb. nov. for a marine bacterium previously
Fitch, W. M. (1971). Toward defining the course of evolution:
assigned to the genus Flectobacillus. Int J Syst Bacteriol 40, 337–347.
minimum changes for a specific tree topology. Syst Zool 20, 406–416.
Saitou, N. & Nei, M. (1987). The neighbor-joining method: a new
Kumar, S., Tamura, K. & Nei, M. (2004). MEGA3: integrated software
method for reconstructing phylogenetic trees. Mol Biol Evol 4, 406–425.
for molecular evolutionary genetics analysis and sequence alignment.
Seidler, R. J. & Mandel, M. (1971). Quantitative aspects of
deoxyribonucleic acid renaturation: base composition, state of
Lane, D. J. (1991). 16S/23S rRNA sequencing. In Nucleic Acid
chromosome replication, and polynucleotide homologies. J Bacteriol
E. Stackebrandt & M. Goodfellow. Chichester: Wiley.
Zhang, D., Yang, H., Huang, Z., Zhang, W. & Liu, S.-J. (2002).
Marmur, J. & Doty, P. (1962). Determination of the base composition
Rhodopseudomonas faecalis sp. nov., a phototrophic bacterium
of deoxyribonucleic acid from thermal denaturation temperature.
isolated from an anaerobic reactor that digests chicken faeces. Int
J Syst Evol Microbiol 52, 2055–2060.
International Journal of Systematic and Evolutionary Microbiology 56
Exploring the Link Between Depression and Early Alzheimer’s Prozac. Zoloft. Paxil. Effexor. Cymbalta. In today’s day and age, I can guarantee you recognize at least one or more of those medications. What do they all have in common you? They are all antidepressants. It is completely understandable for patients with Alzheimer’s disease to become depressed. But evidence suggests that depres
Course Title Lecturer Semester Course Objectives Medical and Pharmaceutical Chemistry (how drugs are made and used) Acquire basic knowledge of pharmaceutical products, such as drug design and how they are developed. In addition, students will deepen their understanding of the use of drugs through learning Course Outline Pharmaceutical products are defined as substances that
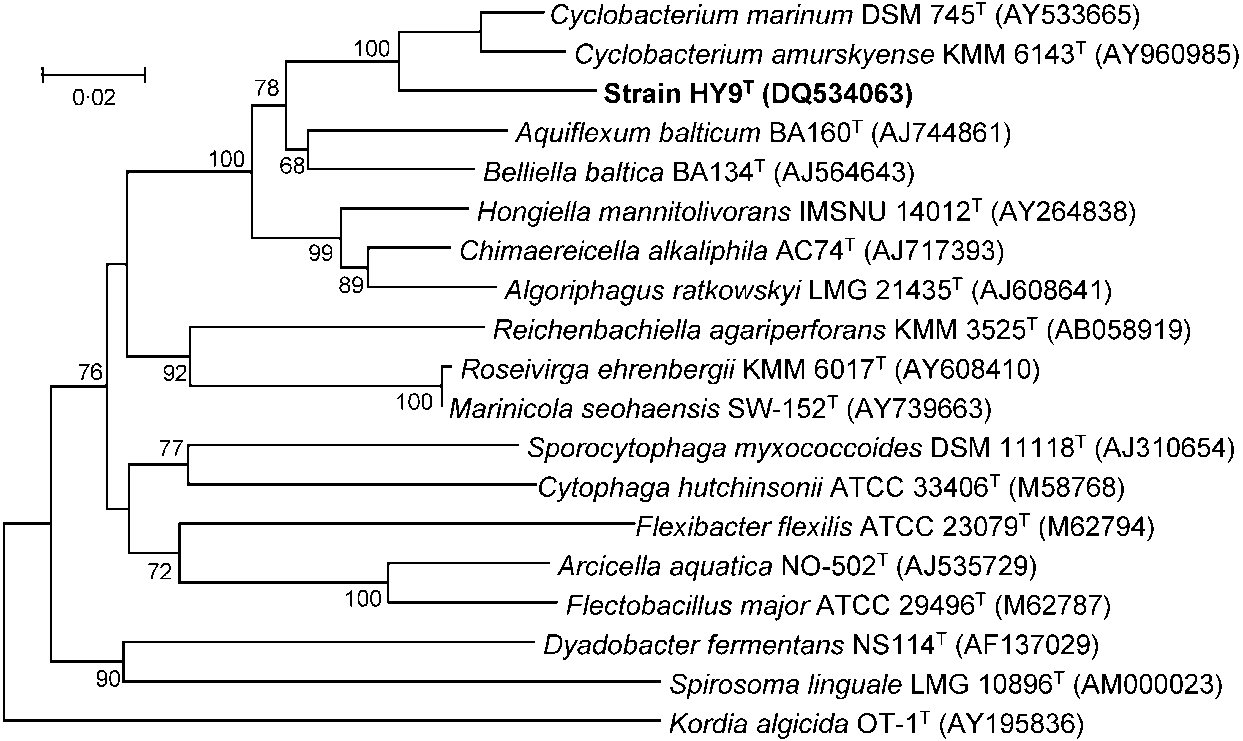 Table 1. Differential properties of strain HY9T and the type
27F and 1492R (Lane, 1991) and sequenced. Preliminary
comparisons with sequences held in GenBank, performedusing BLASTN searches, revealed that the isolate was related
Strains: 1, HY9T; 2, C. marinum LMG 13164T; 3, C. amurskyense
to C. marinum. Further analysis of 16S rRNA gene sequences
KMM 6143T. Data are from Nedashkovskaya et al. (2005) and
was performed using MEGA, version 3.1 (Kumar et al., 2004).
Table 1. Differential properties of strain HY9T and the type
27F and 1492R (Lane, 1991) and sequenced. Preliminary
comparisons with sequences held in GenBank, performedusing BLASTN searches, revealed that the isolate was related
Strains: 1, HY9T; 2, C. marinum LMG 13164T; 3, C. amurskyense
to C. marinum. Further analysis of 16S rRNA gene sequences
KMM 6143T. Data are from Nedashkovskaya et al. (2005) and
was performed using MEGA, version 3.1 (Kumar et al., 2004).