Tadalafil entfaltet seine Wirkung über eine selektive Hemmung der PDE5, wodurch die Konzentration von cGMP im glatten Muskelgewebe stabil bleibt. Diese biochemische Modulation resultiert in einer langanhaltenden Relaxation der Gefäßwände. Der Wirkstoff wird nach oraler Einnahme effizient resorbiert, mit einer Bioverfügbarkeit von rund 80 %. Seine Halbwertszeit von bis zu 36 Stunden ist innerhalb dieser Substanzklasse außergewöhnlich. Abgebaut wird er in der Leber, hauptsächlich durch CYP3A4, mit anschließender biliärer Exkretion. Typische unerwünschte Wirkungen entstehen durch eine verstärkte Vasodilatation, etwa Kopfschmerzen oder Flush. Pharmakologisch wird cialis generika vor allem durch die verlängerte Wirkungsdauer charakterisiert.
Jap december 87/6
Skeletal muscle energy metabolism during prolonged, fatiguing exercise Mark A. Febbraio and Jane Dancey Journal of Applied Physiology 87:2341-2347, 1999. You might find this additional information useful.
This article cites 37 articles, 27 of which you can access free at:
This article has been cited by 2 other HighWire hosted articles:
Evidence for complex system integration and dynamic neural regulation of skeletal muscle recruitment during exercise in humans A St Clair Gibson and T D Noakes
Glycogen availability does not affect the TCA cycle or TAN pools during prolonged, fatiguing exercise J. Baldwin, R. J. Snow, M. J. Gibala, A. Garnham, K. Howarth and M. A. Febbraio
Biochemistry . Polymerase Chain Reaction Biochemistry . Glycogen
Physiology . Exertion Medicine . Exercise Medicine . Fatigue Medicine . Fitness (Physical Activity)
including high-resolution figures, can be found at:
Journal of Applied Physiology can be found at:
This information is current as of April 5, 2005 .
Journal of Applied Physiology publishes original papers that deal with diverse areas of research in applied physiology, especially those papers emphasizing adaptive and integrative mechanisms. It is published 12 times a year (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright 2005 by the American Physiological Society. ISSN: 8750-7587, ESSN: 1522-1601. Visit our website at
Skeletal muscle energy metabolism duringprolonged, fatiguing exercise
MARK A. FEBBRAIO AND JANE DANCEYExercise Physiology and Metabolism Laboratory, Department of Physiology,The University of Melbourne, Parkville, Victoria 3052, AustraliaFebbraio, Mark A., and Jane Dancey. Skeletal muscle
acid cycle intermediates, in turn resulting in an impair-
energy metabolism during prolonged, fatiguing exercise. J.
ment in ATP provision via oxidative phosphorylation
Appl. Physiol. 87(6): 2341–2347, 1999.—A depletion of phos-
(32, 34). Because ATP demand during prolonged exer-
phocreatine (PCr), fall in the total adenine nucleotide pool
cise is maintained, such a decrease in ATP provision
(TAN ϭ ATP ϩ ADP ϩ AMP), and increase in TAN degrada-
leads to transient ADP formation and ATP generation
tion products inosine 5Ј-monophosphate (IMP) and hypoxan-
from alternative pathways, including creatine phospho-
thine are observed at fatigue during prolonged exercise at70% maximal O
kinase (CPK) and adenylate kinase (AK) (32). Because
2 uptake in untrained subjects [J. Baldwin,
R. J. Snow, M. F. Carey, and M. A. Febbraio. Am. J. Physiol.
CPK has a much higher activity in skeletal muscle
277 (Regulatory Integrative Comp. Physiol. 46): R295–R300,
compared with AK (7), phosphocreatine (PCr) has been
1999]. The present study aimed to examine whether these
demonstrated to be an effective buffer of ADP during
metabolic changes are also prevalent when exercise is per-
prolonged exercise until concentrations of PCr are
formed below the blood lactate threshold (LT). Six healthy,
reduced to ϳ40 mmol/kg dry wt (dw), after which time
untrained humans exercised on a cycle ergometer to volun-
AK becomes more active, resulting in a greater forma-
tary exhaustion at an intensity equivalent to 93 Ϯ 3% of LT
tion of AMP, which is rapidly deaminated to inosine
(ϳ65% peak O2 uptake). Muscle biopsy samples were ob-
5Јmonophosphate (IMP) (35). Accordingly, many stud-
tained at rest, at 10 min of exercise, ϳ40 min before fatigue
ies have noted the accumulation of IMP at fatigue
(FϪ40 ϭ143 Ϯ 13 min), and at fatigue (F ϭ 186 Ϯ 31 min). Glycogen concentration progressively declined (P Ͻ 0.01) to
during prolonged exercise in the presence of low intra-
very low levels at fatigue (28 Ϯ 6 mmol glucosyl U/kg dry wt).
muscular glycogen stores (2, 5, 29, 32, 34, 35).
Despite this, PCr content was not different when FϪ40 was
Although these findings suggest an imbalance be-
compared with F and was only reduced by 40% when F was
tween ATP synthesis and degradation rates in the
compared with rest (52.8 Ϯ 3.7 vs. 87.8 Ϯ 2.0 mmol/kg dry wt;
presence of low glycogen stores, it is important to note
P Ͻ 0.01). In addition, TAN concentration was not reduced,
that these studies have been conducted in untrained
IMP did not increase significantly throughout exercise, and
individuals exercising at an intensity of ϳ70% maximal
hypoxanthine was not detected in any muscle samples. A
significant correlation (r ϭ 0.95; P Ͻ 0.05) was observed
˙ O2max). In contrast, recently we (2) and
others (33) have demonstrated that neither the total
between exercise time and glycogen use, indicating that
adenine nucleotide pool (TAN ϭ ATP ϩ ADP ϩ AMP)
glycogen availability is a limiting factor during prolongedexercise below LT. However, because TAN was not reduced,
nor IMP concentrations are significantly changed from
PCr was not depleted, and no correlation was observed
resting values when endurance-trained men exercised
between glycogen content and IMP when glycogen stores
to exhaustion at a similar relative workload despite the
were compromised, fatigue may be related to processes other
presence of low intramuscular glycogen stores. Further-
than those involved in muscle high-energy phosphagen me-
more, Green et al. (23) have observed an elevation in
IMP in the muscles of untrained men during prolonged
total adenine nucleotides; phosphocreatine; lactate thresh-
exercise at a workload corresponding to 70% V
after 30 min of exercise when glycogen stores were notlimited. Importantly, the elevated IMP was not presentat the same time during exercise at the same absoluteworkload after 4 and 8 wk of endurance training. In
IT IS WELL ESTABLISHED that fatigue during prolonged
addition, in our recent study (2) hypoxanthine, an IMP
exercise coincides with low intramuscular glycogen
degradation product that can diffuse from the cell, was
stores (2, 3, 9, 12, 13, 25, 32, 34, 38). Although there are
markedly elevated in plasma after 5 min of exercise,
several possible reasons as to the requirement for
when glycogen stores were unlikely to be compromised,
carbohydrate in the maintenance of contractile force
(for review, see Ref. 18), it is widely accepted that
possible, therefore, that the elevated IMP observed at
metabolic processes are limited by carbohydrate avail-
fatigue in untrained, but not endurance-trained, men
ability. It has been suggested that as muscle glycogen
(2) may occur in the presence of, but may not be caused
stores are progressively compromised during exercise,
by, low intramuscular glycogen stores. In addition,
flux through glycolysis is reduced, leading to a fall in
endurance exercise training also results in attenuated
pyruvate formation and a reduction in tricarboxylic
lactate and ammonia accumulation and PCr degrada-tion (11, 22–24). These findings demonstrate that train-ing improves the match between ATP synthesis and
The costs of publication of this article were defrayed in part by the
degradation during exercise at submaximal work rates.
payment of page charges. The article must therefore be hereby
It is possible, therefore, that the workload of
marked ‘ advertisement’ in accordance with 18 U.S.C. Section 1734
˙ O2max, frequently chosen to examine the relationship
8750-7587/99 $5.00 Copyright 1999 the American Physiological Society
between glycogen availability and muscle energy me-
˙ O2 to verify exercise intensity. Heart
tabolism during fatiguing steady-state exercise, re-
rate was also monitored during this trial via telemetry
quires an ATP turnover rate that cannot be sufficiently
(Sports Tester, Polar). An electric fan was used to circulate air,
met by oxidative metabolism in untrained individuals.
and water was provided ad libitum. Subjects were instructed
The increase in PCr degradation and accumulation of
to cycle at the predetermined work rate, maintaining a pedal
IMP observed in the presence of low glycogen concentra-
frequency of 80–90 rpm until fatigue. Fatigue was defined as
tion may, therefore, be unrelated to glycogen availabil-
the inability to complete one pedal revolution because the
ity but may be due to increased energy provision from
work rate on the electrically braked cycle ergometer wasnon-pedal-frequency dependent. All subjects were given strong
the CPK and AK reactions throughout exercise. Thus
verbal encouragement from the investigators to continue
the purpose of the present study was to examine muscle
energy metabolism in untrained subjects throughout
The experimental trial was conducted at least 7 days after
prolonged, fatiguing exercise at a workload where the
the familiarization trial. The protocol was identical to the
ATP demand was met via oxidative sources. We hypoth-
familiarization trial but included venous blood and muscle
esized that although glycogen would be depleted at
sampling at various stages throughout exercise. In addition
fatigue, there would be little, if any, disruption to the
˙ O2 measurements, pulmonary gases were analyzed for the
respiratory exchange ratio during this trial.
Venous blood samples were obtained by using a 20-gauge
Teflon catheter (Terumo, Tokyo, Japan) inserted into a vein inthe antecubital space. The vein was kept patent by flushing
Subjects. Six healthy but untrained subjects [20.7 Ϯ 2.4 yr;
with 0.5 ml sodium chloride-5 U heparin after each sample
˙ O2peak) ϭ 2.49 Ϯ 0.5 l/min] collection. Muscle samples were obtained from the vastus
volunteered for the experiment. The experimental procedures
lateralis by using the percutaneous needle biopsy technique
and possible risks of the study were explained to all subjects
with suction. Briefly, local anesthetic was injected ϳ10 cm
before they gave their informed, written consent. The experi-
and 13 cm proximal to the lateral epicondyle of the femur of
ment was approved by the Human Research Ethics Commit-
both legs. Four separate incisions (2 in each leg) were then
made over the anesthetized areas, and muscle samples were
˙ O2peak and lactate threshold (LT) determination. Each obtained at rest, at 10 min of exercise (10 min), ϳ40 min
subject initially performed a cycling test to volitional fatigueon an electromagnetically braked cycle ergometer (Lode
before fatigue (FϪ40 ϭ 143 Ϯ 14 min), and at fatigue (F ϭ
Instrument, Groningen, The Netherlands) to determine
186 Ϯ 31 min). FϪ40 was estimated from the results obtained
during the familiarization trial. On sampling, the muscle was
O2peak and LT. Expired air was directed into Douglas bags
via a Hans Rudolf valve and plastic tubing. Oxygen and
rapidly frozen in liquid nitrogen for later metabolite analysis.
carbon dioxide content of the Douglas bags were analyzed by
The time from the cessation of exercise to freezing was ϳ10 s.
using Applied Electrochemistry (Ametek, Pittsburgh, PA)
Tissue treatment and analysis. After each blood sample
S-3A/II and CD-3A gas analyzers, calibrated before each test
collection, blood was placed in fluoride heparin, mixed, and
with a commercially prepared gas mixture of known composi-
spun for 3 min at 8,000 rpm. The plasma supernatant was
tion. The volumes of expired gases were determined by using
then removed, stored on ice until completion of the trial, and
a gas meter (Parkinson-Cowan, Manchester, UK). V
˙ O2peak was then frozen until later analysis of plasma glucose and lactate
calculated by using standard equations (8). During this test,
by using an automated method (EML-105, Electrolyte Metabo-
venous blood samples were also obtained at rest and at the
lite Laboratory, Radiometer, Copenhagen, Denmark). A fur-
completion of every increment in the workload. Samples of
ther 1.5 ml of whole blood were placed in tubes containing 30
whole blood were immediately mixed in a tube containing
µl of EGTA/GSH. This tube was placed on ice until the
lithium heparin. A 125-µl aliquot of whole blood was added to
completion of the trial and spun as previously described. The
250 µl perchloric acid and spun in a centrifuge, and the
plasma was then frozen for later analysis of free fatty acids by
supernatent was frozen and stored for subsequent lactate
using an enzymatic colorimetric method (Nefa-C kit, Wako
determination (26). Each subject’s LT was determined accord-
Pure Chemicals) according to the methods of Miles et al. (27).
ing to the methods of Coyle et al. (14). Briefly, the increase in
Muscle samples were freeze-dried for 24 h, dissected free of
blood lactate was plotted against O2 uptake (V
˙ O2). After any blood and connective tissue, powdered, extracted, and
determination of the lactate steady state during the initial
analyzed for glycogen, lactate, adenine nucleotides (ATP,
incremental workloads, a value corresponding to 1 mmol/l
ADP, AMP) and their degradation products (IMP and hypoxan-
above this point was taken as the LT. The corresponding V
˙ O2 thine), PCr, and creatine (Cr) as previously described (17).
at this point was multiplied by 0.95 to calculate 95% LT. The
The concentrations of ATP, ADP, AMP, IMP, PCr, and Cr were
desired workload was then determined from the V
˙ O2 vs. adjusted to the peak total PCrϩCr concentration for each
Experimental procedure. At least 1 wk after the V
subject. This procedure minimized the error in measuring
test, subjects returned to the laboratory to perform a familiar-
nonmuscle components of the tissue not visible in the sample.
ization trial. This trial served to familiarize subjects with the
Lactate and glucose were not corrected because of their
cycling protocol and enabled us to check the workload and
determine an approximate time to fatigue. Subjects were
Statistical analyses. A one-way ANOVA with repeated
instructed to refrain from alcohol, caffeine, tobacco, and
measures on the time factor was used to compare blood and
strenuous exercise and to consume their normal diet for the
muscle metabolite data throughout the trial. A Newman-
preceding 24 h. Subjects arrived in the laboratory in the
Keuls post hoc test was used to locate difference when the
morning after an overnight fast, were weighed, and then
ANOVA revealed a significant interaction. Correlation coeffi-
commenced cycling on the previously mentioned cycle ergom-
cients were determined by using Pearson’s product moments.
eter at the predetermined workload. Expired gases were
All data are reported as means Ϯ SE unless otherwise stated.
collected via Douglas bags during this trial and analyzed at
The level of significance for all tests was set at P Ͻ 0.05.
Subjects cycled for 186 Ϯ 31 min at a workload that
corresponded to 93 Ϯ 8% LT. This was equivalent to1.60 Ϯ 0.1 l/min or ϳ64% V
plasma lactate accumulation increased (P Ͻ 0.05) inthe initial period of exercise, but concentrations re-turned to resting levels thereafter, indicating that thecontribution to energy metabolism via anaerobic glycoly-sis was minimal (Fig. 1).
Heart rate initially increased (P Ͻ 0.05) but reached
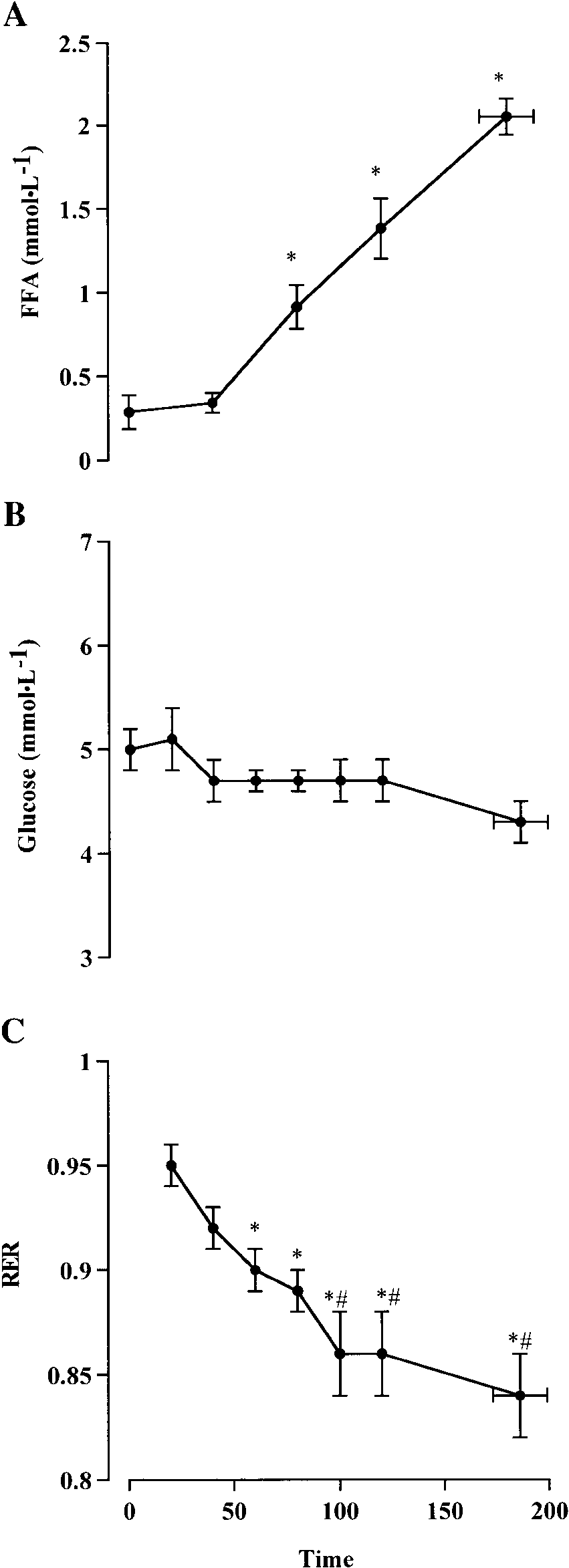
throughout exercise (data not shown). Although therespiratory exchange ratio progressively fell (P Ͻ 0.05)during the first 80 min, it was maintained thereafter(Fig. 2). In addition, plasma glucose concentration didnot alter throughout exercise, indicating that circulat-ing glucose availability was not compromised at fatigue
Fig. 2. Plasma free fatty acids (FFA; A), plasma glucose (B), andrespiratory exchange ratio (RER; C) throughout cycling exercise tofatigue, at 93 Ϯ 8% of lactate threshold. Values are means Ϯ SE; n ϭ 6subjects. *Different (P Ͻ 0.05) from R. #Different (P Ͻ 0.5) from 80 min.
Fig. 1. Muscle (A) and plasma (B) lactate concentrations at rest andthroughout cycling exercise to fatigue, at 93 Ϯ 8% of lactate thresh-
(Fig. 2). Plasma free fatty acid concentrations increased
old. Muscle lactate measurements were taken at rest (R), 10 min of
(P Ͻ 0.05) after 60 min of exercise (Fig. 2).
exercise (10), 40 min before fatigue (FϪ40), and fatigue (F). Plasma
Muscle glycogen concentration decreased (P Ͻ 0.05)
lactate measurements were taken at rest (0 min) and every 20 min
progressively throughout exercise, and concentrations
throughout exercise. Values are means Ϯ SE; n ϭ 6 subjects. * Different (P Ͻ 0.05) from rest.
were very low (Ͻ50 mmol glucosyl U/kg dw) at F (Fig.
Fig. 3. Muscle glycogen at R, 10, FϪ40, and F during cycling exerciseat 93 Ϯ 8% of lactate threshold. Values are means Ϯ SE; n ϭ 6subjects. * Different (P Ͻ 0.05) from R. # Different (P Ͻ 0.05) from 10.
3). However, despite the fact that glycogen content was
decreased by 50% in all subjects when FϪ40 wascompared with F, this decrease was not statisticallysignificant. Muscle PCr was higher (P Ͻ 0.05) whenrest was compared with 10 min, FϪ40, and F. Of note isthe fact that, although PCr declined (P Ͻ 0.05) asexercise progressed beyond 10 min, it did not decreasewhen FϪ40 was compared with F (Table 1). Conversely,
the concentration of intramuscular Cr increased (P Ͻ0.05) throughout exercise and was different from rest at10 min, FϪ40, and F (Table 1). There was no change inmuscle ATP, ADP, or AMP concentrations throughoutexercise, and, therefore, the TAN pool remained un-changed throughout exercise (Table 1). Although thereappeared to be a tendency for IMP to accumulate
throughout exercise, this did not reach statistical signifi-cance (P Ͼ 0.05) (Table 1). Hypoxanthine was notdetected in any sample despite an analytic detectionlimit of between 0.005 and 0.01 mmol/kg dw. Further-more, the change in IMP throughout exercise was notdifferent (Fig. 4), and there was no correlation (r ϭ
Fig. 4. Change in inosine 5Ј-monophosphate (IMP) from rest to 10
0.056, P Ͼ 0.05) between IMP and glycogen content at
min of exercise (10–0 min), from 10 min of exercise to ϳ40 min before
either FϪ40 (r ϭ 0.083; P Ͼ 0.05) or F (r ϭ 0.73; P Ͼ
fatigue (FϪ40–10), and from fatigue to ϳ40 min before fatigue(F–FϪ40) during cycling exercise at 93 Ϯ 8% of lactate threshold (A)and relationship between IMP and glycogen at FϪ40 (r) and at F (s;
Table 1. Muscle metabolite concentrations beforeB). Values are means Ϯ SE; n ϭ 6 subjects. exercise, at 10 min of exercise, at 40 min beforefatigue, and at fatigue during cycling exercise
0.05) (Fig. 4). In contrast, a significant correlation (r ϭ
at 93 Ϯ 8% of blood lactate threshold
0.95; P Ͻ 0.05) was observed between time to exhaus-tion and glycogen use (Fig. 5). DISCUSSION
This study is the first to measure muscle energy
metabolism throughout exercise in untrained subjects
at a workload below the LT, where ATP supply from
oxidative metabolism is sufficient to meet the ATP
demand. Unlike previous studies conducted in un-
Values are means Ϯ SE in mmol/kg dry wt; n ϭ 6 subjects. Rest,
trained subjects (2, 5, 32, 34), the results from this
before exercise; 10 min, 10 min of exercise; F Ϫ 40, 40 min before
study suggest that despite compromised intramuscular
fatigue (F); PCr, phosphocreatine; Cr, creatine; TAN, total adenine
glycogen stores, fatigue appears to be associated with
nucleotide pool (TANϭATPϩADPϩAMP); IMP, inosine 5Ј-monophos-
factors other than those related to muscle high-energy
phate. * Different (P Ͻ 0.05) from rest. † Different (P Ͻ 0.05) from 10min.
phosphagen metabolism. The relationship between gly-
˙ O2max, factors such as metabolic acido-
sis may have contributed to fatigue before glycogendepletion.
It is important to note, however, that although
glycogen was reduced by ϳ50% in all subjects whenFϪ40 is compared with F, the fall was not statisticallysignificant. Although unlikely, because of the relation-ship between glycogen use and exercise duration (Fig. 5), the possibility cannot be ruled out that fatigue wasrelated to factors other than glycogen availability, suchas a decrease in the central drive to exercise. It hasbeen proposed for a number of years (39) that theserotoninergic system may play a crucial role in thecentral control of fatigue during prolonged exercise. Inaddition, prolactin has been proposed as a marker ofcentral serotoninergic activity, and increases in plasmaprolactin concentration have been observed as exerciseintensity increased (16). Recent evidence suggests thatmuscular contraction increases reactive oxygen speciesin skeletal muscle, which promote low-frequency fa-
Fig. 5. Correlation between exercise duration and glycogen utiliza-
tigue in vitro (30). Therefore, the impairment of contrac-
tion during cycling exercise at 93 Ϯ 8% of lactate threshold. Values
tile function may be independent of glycogen availabil-
are means Ϯ SE; n ϭ 6 subjects.
ity. Further investigations into the role of the centralnervous system and reactive oxygen species production
cogen use and exercise duration (Fig. 5), as well as the
during prolonged exercise to fatigue are needed.
very low levels of glycogen within the muscles at
In the present study, TAN did not fall, IMP did not
fatigue, supports previous studies (2, 3, 9, 12, 13, 25, 32,
significantly accumulate, PCr was not reduced when F
34, 38) that suggest that glycogen availability may be a
was compared with FϪ40 (Table 1), and no hypoxan-
limiting factor during steady-state exercise. However,
thine was detected in the muscle samples. Taken together,
because TAN was not reduced, PCr degradation did not
these data demonstrate that the intracellular high-energy
fall at fatigue, IMP did not significantly accumulate
phosphagen pool was not affected to a great extent. Even
(Table 1), and no correlation was observed between
though there was a tendency for IMP to accumulate,
glycogen content and IMP late in exercise (Fig. 4), there
the important factor was that there was no correlation
was little evidence that this reduced glycogen availabil-
between IMP accumulation and glycogen concentration
ity had a major influence on muscle high-energy phos-
near the end of exercise, when glycogen levels were
compromised. In fact, the subject with the lowest
The workload selected in the present study was
muscle glycogen content at fatigue displayed no detect-
designed such that the ATP requirement was ad-
able IMP accumulation, whereas the subject with the
equately met by oxidative processes. Although lactate
highest glycogen at fatigue had the highest IMP level at
concentration increased in both muscle and plasma at
this point (Fig. 4). This lack of a correlation between
the onset of exercise, this rise was transient, and
IMP and glycogen content when glycogen is compro-
concentration fell to resting levels thereafter (Fig. 1).
mised is in contrast to the data of Sahlin et al. (33). Of
Although these data only reflect a balance between
note, however, is the fact that in the previous study the
lactate production and removal, they suggest that ATP
exercise intensity at which the subjects exercised ranged
supply from oxidative metabolism was sufficient in
meeting energy demand. Given this, it was not surpris-
correlation was significant, three of seven subjects
ing that these untrained subjects were able to exercise
demonstrated no IMP accumulation at fatigue.
for ϳ3 h. In addition, the low muscle glycogen levels
Another important finding in this study was that the
observed at fatigue were expected because carbohy-
small accumulation of IMP occurred progressively
drate has been demonstrated to provide ϳ50% of the
throughout exercise and did not occur late in exercise,
total energy metabolized during exercise at this inten-
when glycogen was compromised (Fig. 4). In fact, when
sity (31). Interestingly, the concentration of glycogen at
a power analysis was performed on these data, the
FϪ40 was lower than that previously observed at
number of subjects needed to obtain a statistical differ-
fatigue in some studies (2, 32, 35). This is probably due
ence was 102. Therefore, despite the facts that IMP
to the important fact that this is the only study to date
rose slightly over time and our subject number was low,
that has normalized the exercise intensity to a marker
we are confident that our data demonstrate no biologi-
of metabolic stress rather than to a percentage of
cal relationship between glycogen content and IMP
˙ O2max. Therefore, factors such as metabolic acidosis, formation. In addition, the fact that the TAN pool was
which can disrupt contractile processes, could not have
not altered at all suggests that the small and insignifi-
resulted in fatigue in the present study. In contrast, in
cant rise in IMP over time was physiologically unimpor-
previous studies where the workload was normalized to
A limitation of the present study is that analyses
lar processes, which may cause a disturbance in contrac-
were conducted on whole, mixed-fiber muscle samples.
It is possible that the accumulation of IMP may havebeen related to fiber-type activation. As discussed above,
The authors acknowledge the technical assistance of Jo Ann
Parkin, Damien Angus, and Kirsten Howlett, and the medical
IMP accumulated progressively throughout exercise
assistance of Dr. Andrew Garnham. The authors also acknowledge
rather than at fatigue (Fig. 4). It has been demon-
Dr. Michael Carey for generous use of his laboratory in the HPLC
strated that type II fiber activity increases as submaxi-
analyses and Dr. Rod Snow for assistance in preparing this manu-
mal-intensity exercise progresses (20). Furthermore,
script. We also thank the subjects for their participation in this study.
Address for reprint requests and other correspondence: M. A.
Norman et al. (28) demonstrated that glycogen-de-
Febbraio, Dept. of Physiology, The Univ. of Melbourne, Parkville,
pleted type II fibers accumulate more IMP compared
Victoria 3052, Australia (E-mail: m.febbraio@physiology.unimelb.
with type I fibers. It would have been desirable in the
present study to perform pooled single-fiber analyses.
Received 19 May 1999; accepted in final form 16 August 1999.
However, given the small content of IMP in mixedmuscle and the present analytic techniques, this was
REFERENCES
1. Angus, D. J., M. A. Febbraio, and M. Hargreaves. Glucose
It is important to note that as intramuscular glyco-
kinetics during prolonged exercise to fatigue with carbohydrate
gen stores became compromised toward the end of
ingestion (Abstract). Med. Sci. Sport. Exer. 31: S127, 1999.
exercise in the present study, circulating glucose did
2. Baldwin, J., R. J. Snow, M. F. Carey, and M. A. Febbraio.
Muscle IMP accumulation during fatiguing submaximal cycling
not fall (Fig. 3). In fact, two subjects were relatively
in endurance trained and untrained men. Am. J. Physiol. 277
hyperglycemic at fatigue when compared with rest,
(Regulatory Integrative Comp. Physiol. 46): R295–R300, 1999.
despite glycogen levels being Ͻ50 mmol/kg dw in all
3. Bergstrom, J., L. Hermansen, E. Hultman, and B. Saltin.
subjects. This is in agreement with some (2, 34, 37, 38),
Diet, muscle glycogen and physical performance. Acta Physiol. Scand. 71: 140–150, 1967.
but not all (9, 13), previous studies. Although, there are
4. Booth, J., M. J. McKenna, P. A. Ruell, T. H. Gwinn, G. M.
no published data to our knowledge that examine
Davis, M. W. Thompson, A. R. Harmer, S. K. Hunter, and
glucose uptake during prolonged exercise to fatigue, we
J. R. Sutton. Impaired calcium pump function does not slow
have recently demonstrated that when euglycemia is
relaxation in human skeletal muscle after prolonged exercise. J. Appl. Physiol. 83: 511–521, 1997.
maintained during prolonged exercise, isotopic-tracer-
5. Broberg, S., and K. Sahlin. Adenine nucleotide degradation in
determined glucose uptake (rate of disappearance)
human skeletal muscle during prolonged exercise. J. Appl.
neither falls nor indeed plateaus at the point of fatigue
Physiol. 67: 116–122, 1989.
(1). This observation is important when the effect of
6. Byrd, S. K., A. K. Bode, and G. A. Klug. Effects of exercise of
varied duration on sarcoplasmic reticulum function. J. Appl.
carbohydrate availability on muscle energy metabolism
Physiol. 66: 1383–1389, 1989.
is considered. If glucose availability, glucose rate of
7. Chi, M. M.-Y., C. S. Hintz, E. F. Coyle, W. H. Martin III, J. L.
disappearance, and carbohydrate oxidation are not
Ivy, P. M. Nemeth, J. O. Holloszy, and O. H. Lowry. Effect of detraining on enzymes of energy metabolism in individual
compromised, one would expect exercise to continue if
human muscle fibers. Am. J. Physiol. 244 (Cell Physiol. 13):
fatigue is related to muscle energy metabolism because
glucose moieties would be available for flux through
8. Chin, E. R., and D. G. Allen. Effects of reduced muscle glycogen
concentration on force, Ca2ϩ release and contractile proteinfunction in intact mouse skeletal muscle. J. Physiol. (Lond.) 498:
Although there was little evidence of metabolic stress
within the muscle at fatigue, the relationship between
9. Coggan, A. R., and E. F. Coyle. Reversal of fatigue during
glycogen content and exercise duration suggests that
prolonged exercise by carbohydrate infusion or ingestion. J.
the maintenance of contractile force is dependent on
Appl. Physiol. 63: 2388–2395, 1987.
10. Consolazio, C. G., R. E. Johnson, and L. J. Pecora. Physiologi-
glycogen availability. It has been previously suggested
cal Measurements of Metabolic Functions in Man. New York:
that glycogen may be required for contractile processes
independent of energy metabolism (21). Studies in both
11. Constable, S. H., R. J. Favier, J. A. McLane, R. D. Fell, M.
animals (6, 8, 36) and humans (4) have suggested a link
Chen, and J. O. Holloszy. Energy metabolism in contracting rat skeletal muscle: adaptation to exercise training. Am. J.
between sarcoplasmic reticulum Ca2ϩ uptake and re-
Physiol. 253 (Cell Physiol. 22): C316–C322, 1987.
lease and glycogen availability. In addition, topographi-
12. Constantin-Teodosiu, D., G. Cederblad, and E. Hultman.
cal localization of glycogen within human skeletal
PDC activity and acetyl group accumulation in skeletal muscle
muscle has been observed (19). Therefore, depletion of
during prolonged exercise. J. Appl. Physiol. 73: 2403–2407, 1992.
glycogen in the sarcoplasmic reticulum may possibly
13. Coyle, E. F., A. R. Coggan, M. K. Hemmert, and J. L. Ivy.
Muscle glycogen utilization during prolonged strenuous exercise
lead to a failure of contractile force, although further
when fed carbohydrate. J. Appl. Physiol. 61: 165–172, 1986.
14. Coyle, E. F., W. H. Martin, A. A. Ehsani, J. M. Hagberg, S. A.
In summary, the data from this study indicate that
Bloomfield, D. R. Sinacore, and J. O. Holloszy. Blood lactate
fatigue during prolonged exercise may be related to
threshold in some well-trained ischemic heart disease patients. J. Appl. Physiol. 54: 18–23, 1983.
carbohydrate availability. It is clear, however, that
15. Davis, J. M., S. P. Bailey, J. A. Woods, F. J. Galiano, M. T.
when untrained subjects exercise below their LT, there
Hamilton, and W. P. Bartoli. Effects of carbohydrate feedings
is little evidence of compromised high-energy metabo-
on plasma free tryptophan and branched chain amino acids
lism within the contracting muscle at fatigue. It is
during prolonged cycling. Eur. J. Appl. Physiol. 65: 513–519,1992.
possible, therefore, that in these circumstances insuffi-
16. DeMeirleir, K. L., L. Baeyens, M. L’Hermite-Baleriaux, M.
cient carbohydrate availability may affect other cellu-
L’Hermite, and W. Hollmann. Exercise-induced prolactin re-
lease is related to anaerobiosis. J. Clin. Endocrinol. Metab. 60:
29. Norman, B., A. Sollevi, L. Kaijser, and E. Jansson. ATP
breakdown products in human muscle during prolonged exercise
17. Febbraio, M. A., R. J. Snow, C. G. Stathis, M. Hargreaves,
to exhaustion. Clin. Physiol. 7: 503–509, 1987. and M. F. Carey. Effect of heat stress on muscle energy
30. Reid, M. B., K. E. Haak, K. M. Franchek, P. A. Valberg, L.
metabolism during exercise. J. Appl. Physiol. 77: 2827–2831,
Kobzik, and M. S. West. Reactive oxygen in skeletal muscle. I.
Intracellular oxidant kinetics and fatigue in vitro. J. Appl.
18. Fitts, R. H. Cellular mechanisms of muscular fatigue. Physiol. Physiol. 73: 1797–1804, 1992.
31. Romijn, J. A., E. F. Coyle, L. S. Sidossis, A. Gastaldelli, J. F. Horowitz, E. Endert, and R. R. Wolfe. Regulation of endog-
19. Friden, J., J. Seger, and B. Eckblom. Topographical localiza-
enous fat and carbohydrate metabolism in relation to exercise
tion of muscle glycogen: an ultrahistochemical study in the
intensity and duration. Am. J. Physiol. (Regulatory Integrative
human vastus lateralis. Acta Physiol. Scand. 135: 381–391,
Comp. Physiol. 34) 265: E380–E391, 1993.
32. Sahlin, K., A. Katz, and S. Broberg. Tricarboxylic acid cycle
20. Gollnick, P. D., R. B. Armstrong, C. W. Saubert IV, W. L.
intermediates in human muscle during prolonged exercise. Am. J.Sembrowich, R. E. Shepherd, and B. Saltin. Glycogen deple- Physiol. 259 (Cell Physiol. 28): C834–C841, 1990.
tion patterns in human skeletal muscle fibres during prolonged
33. Sahlin, K., K. So ¨ derlund, M. Tonkonogi, and K. Hirakoba. ¨ gers Arch. 344: 1–12, 1973.
Phosphocreatine content in single fibers of human muscle after
21. Green, H. J. How important is endogenous muscle glycogen to
sustained submaximal exercise. Am. J. Physiol. 273 (Cell Physiol.
fatigue in prolonged exercise? Can. J. Physiol. Pharmacol. 69:
34. Spencer, M. K., Z. Yan, and A. Katz. Carbohydrate supplemen-
22. Green, H. J., R. Helyar, M. Ball-Burnett, N. Kowalchuk, S.
tation attenuates IMP accumulation in human muscle during
Symon, and B. Farrance. Metabolic adaptations to training
prolonged exercise. Am. J. Physiol. 261 (Cell Physiol. 30): C71–
precede changes in mitochondrial capacity. J. Appl. Physiol. 72:
35. Spencer, M. K., Z. Yan, and A. Katz. Effect of low glycogen on
23. Green, H. J., S. Jones, M. Ball-Burnett, B. Farrance, and D.
carbohydrate and energy metabolism in human muscle during
Ranney. Adaptations in muscle metabolism to prolonged volun-
exercise. Am. J. Physiol. 262 (Cell Physiol. 31): C975–C979,
tary exercise and training. J. Appl. Physiol. 78: 138–145, 1995.
24. Green, H. J., S. Jones, M. E. Ball-Burnett, D. Smith, J.
36. Stephenson, G. D., L. T. Nguyen, and G. M. M. Stephenson.
Glycogen content and excitation-contraction coupling in mechani-
Livesay, and B. W. Farrance. Early muscular and metabolic
cally skinned muscle fibres of the cane toad. J. Physiol. (Lond.).
adaptations to prolonged exercise training in humans. J. Appl.Physiol. 70: 2032–2038, 1991.
37. Thomas, D. E., J. R. Brotherhood, and J. C. Brand. Carbohy-
25. Hermansen, L., E. Hultman, and B. Saltin. Muscle glycogen
drate feeding before exercise: effect of the glycemic index. Int. J.
during prolonged severe exercise. Acta Physiol. Scand. 71: 129–
Sports Med. 12: 180–186, 1991.
38. Tsintzas, O.-K., C. Williams, L. Boobis, and P. Greenhaff.
26. Lowry, O., and J. Passonneau. A Flexible System of Enzymatic
Carbohydrate ingestion and single muscle fiber glycogen metabo-
Analysis. New York: Academic, 1972.
lism during prolonged running in men. J. Appl. Physiol. 81:
27. Miles, J., R. Glassock, J. Aikens, J. Gerich, and M. Hay- mond. A microfluorometric method for the determination of free
39. Wilson, W. M., and R. J. Maughan. Evidence for a possible role
fatty acids in plasma. J. Lipid Res. 24: 96–99, 1983.
of 5-hydroxytryptamine in the genesis of fatigue in man: admin-
28. Norman, B., A. Sollevi, and E. Jansson. Increased IMP
istration of paroxetine, a 5-HT re-uptake inhibitor, reduces the
content in glycogen-depleted muscle fibres during submaximal
capacity to perform prolonged exercise. Exp. Physiol. 77: 921–
exercise in man. Acta Physiol. Scand. 133: 97–100, 1988.
Embryonic muscle development in fast‐ and slow‐growing chickens: role for IGF‐I and myogenic transcription factors Sara L. Al-Musawi and Neil C. Stickland Department of Veterinary Basic Sciences, The Royal Veterinary College, Royal College Street, London NW1 0TU, UK. Introduction Results: embryonic movement Results: gene expression anal
SECONDA UNIVERSITÁ DEGLI STUDI DI NAPOLI FACOLTÁ DI MEDICINA E CHIRURGIA DIPARTIMENTO DI PSICHIATRIA, NEUROPSICHIATRIA INFANTILE, AUDIOFONIATRIA E DERMATOVENEREOLOGIA Direttore: Prof. Mario Maj IL DISTURBO POST-TRAUMATICO DA STRESS: VALUTAZIONE DELL’EFFICACIA DELL’EYE MOVEMENT DESENSITIZATION AND REPROCESSING IL DISTURBO POST- TRAUMATICO DA STRESS
 Skeletal muscle energy metabolism during prolonged,
Skeletal muscle energy metabolism during prolonged,
 Skeletal muscle energy metabolism duringprolonged, fatiguing exercise
MARK A. FEBBRAIO AND JANE DANCEYExercise Physiology and Metabolism Laboratory, Department of Physiology,The University of Melbourne, Parkville, Victoria 3052, Australia
Febbraio, Mark A., and Jane Dancey. Skeletal muscle
Skeletal muscle energy metabolism duringprolonged, fatiguing exercise
MARK A. FEBBRAIO AND JANE DANCEYExercise Physiology and Metabolism Laboratory, Department of Physiology,The University of Melbourne, Parkville, Victoria 3052, Australia
Febbraio, Mark A., and Jane Dancey. Skeletal muscle between glycogen availability and muscle energy me-
˙ O2 to verify exercise intensity. Heart
tabolism during fatiguing steady-state exercise, re-
rate was also monitored during this trial via telemetry
quires an ATP turnover rate that cannot be sufficiently
(Sports Tester, Polar). An electric fan was used to circulate air,
met by oxidative metabolism in untrained individuals.
between glycogen availability and muscle energy me-
˙ O2 to verify exercise intensity. Heart
tabolism during fatiguing steady-state exercise, re-
rate was also monitored during this trial via telemetry
quires an ATP turnover rate that cannot be sufficiently
(Sports Tester, Polar). An electric fan was used to circulate air,
met by oxidative metabolism in untrained individuals.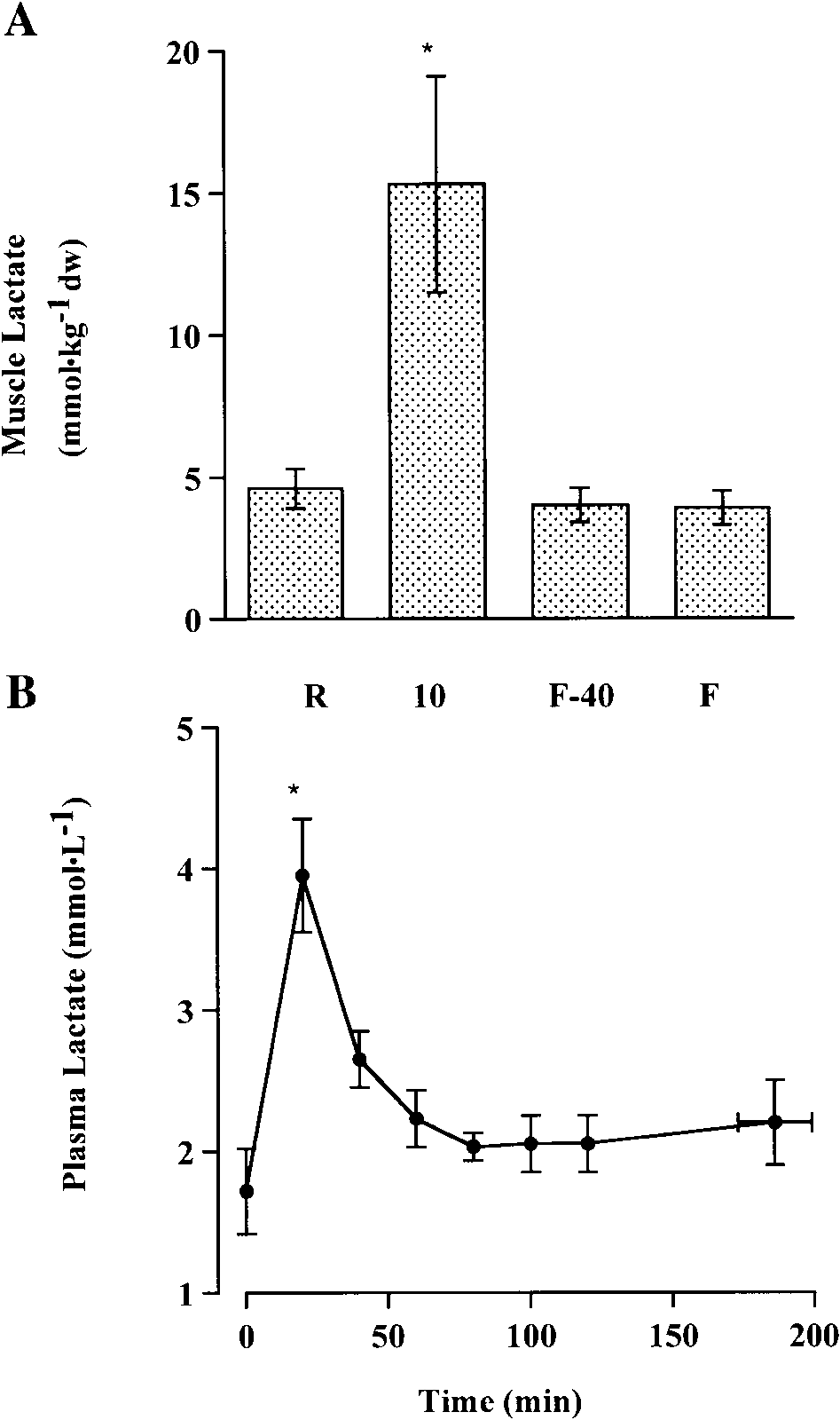
 Subjects cycled for 186 Ϯ 31 min at a workload that
corresponded to 93 Ϯ 8% LT. This was equivalent to1.60 Ϯ 0.1 l/min or ϳ64% V
plasma lactate accumulation increased (P Ͻ 0.05) inthe initial period of exercise, but concentrations re-turned to resting levels thereafter, indicating that thecontribution to energy metabolism via anaerobic glycoly-sis was minimal (Fig. 1).
Subjects cycled for 186 Ϯ 31 min at a workload that
corresponded to 93 Ϯ 8% LT. This was equivalent to1.60 Ϯ 0.1 l/min or ϳ64% V
plasma lactate accumulation increased (P Ͻ 0.05) inthe initial period of exercise, but concentrations re-turned to resting levels thereafter, indicating that thecontribution to energy metabolism via anaerobic glycoly-sis was minimal (Fig. 1).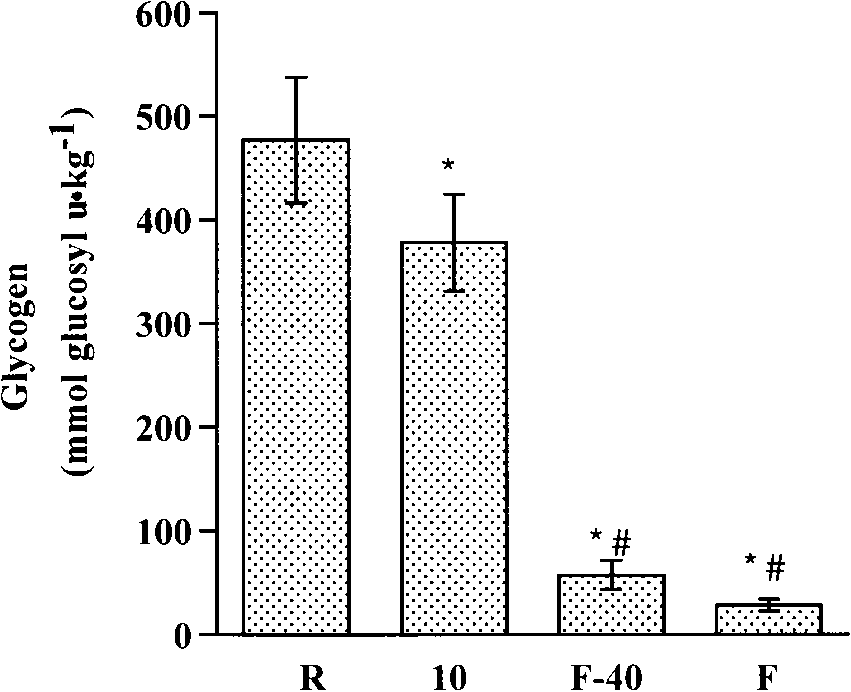
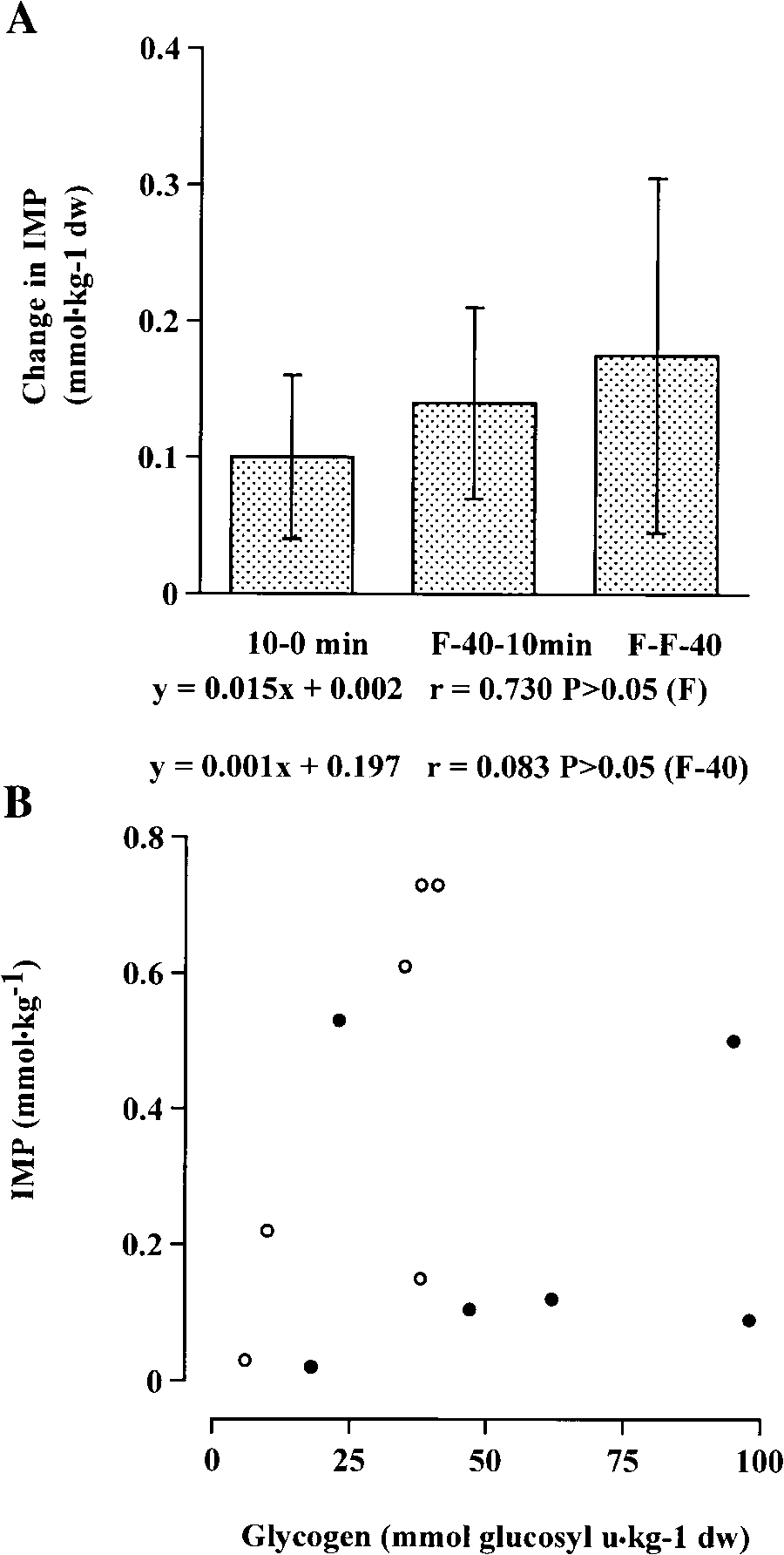
 Fig. 3. Muscle glycogen at R, 10, FϪ40, and F during cycling exerciseat 93 Ϯ 8% of lactate threshold. Values are means Ϯ SE; n ϭ 6subjects. * Different (P Ͻ 0.05) from R. # Different (P Ͻ 0.05) from 10.
Fig. 3. Muscle glycogen at R, 10, FϪ40, and F during cycling exerciseat 93 Ϯ 8% of lactate threshold. Values are means Ϯ SE; n ϭ 6subjects. * Different (P Ͻ 0.05) from R. # Different (P Ͻ 0.05) from 10.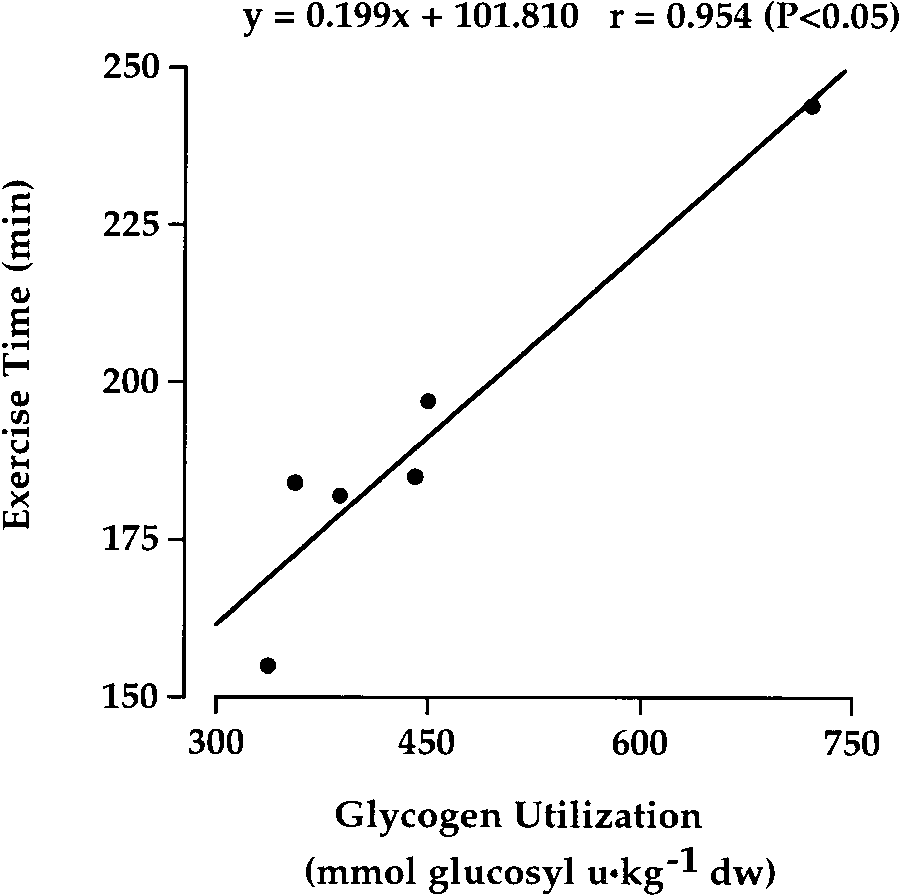
 ˙ O2max, factors such as metabolic acido-
sis may have contributed to fatigue before glycogendepletion.
˙ O2max, factors such as metabolic acido-
sis may have contributed to fatigue before glycogendepletion. A limitation of the present study is that analyses
lar processes, which may cause a disturbance in contrac-
were conducted on whole, mixed-fiber muscle samples.
A limitation of the present study is that analyses
lar processes, which may cause a disturbance in contrac-
were conducted on whole, mixed-fiber muscle samples. lease is related to anaerobiosis. J. Clin. Endocrinol. Metab. 60:
29. Norman, B., A. Sollevi, L. Kaijser, and E. Jansson. ATP
lease is related to anaerobiosis. J. Clin. Endocrinol. Metab. 60:
29. Norman, B., A. Sollevi, L. Kaijser, and E. Jansson. ATP