Tadalafil entfaltet seine Wirkung über eine selektive Hemmung der PDE5, wodurch die Konzentration von cGMP im glatten Muskelgewebe stabil bleibt. Diese biochemische Modulation resultiert in einer langanhaltenden Relaxation der Gefäßwände. Der Wirkstoff wird nach oraler Einnahme effizient resorbiert, mit einer Bioverfügbarkeit von rund 80 %. Seine Halbwertszeit von bis zu 36 Stunden ist innerhalb dieser Substanzklasse außergewöhnlich. Abgebaut wird er in der Leber, hauptsächlich durch CYP3A4, mit anschließender biliärer Exkretion. Typische unerwünschte Wirkungen entstehen durch eine verstärkte Vasodilatation, etwa Kopfschmerzen oder Flush. Pharmakologisch wird cialis generika vor allem durch die verlängerte Wirkungsdauer charakterisiert.
Vimp-06.qxd
Viral Ion Channels/viroporins Viral Proteins that Enhance Membrane Permeability María Eugenia González and Luis Carrasco 1. Introduction
During the infection of cells by animal viruses, membrane permeability is modified at
two different steps of the virus life cycle (Carrasco, 1995) (Figure 6.1). Initially, when thevirion enters cells, a number of different-sized molecules are able to co-enter the cytoplasmwith the virus particles (Fernandez-Puentes and Carrasco, 1980; Otero and Carrasco, 1987). Membrane potential is reversibly destroyed, being restored several minutes later. Endosomesare involved in the co-entry process, since inhibitors of the proton ATPase block early per-meabilization even with viruses that do not require endosomal function. A chemiosmoticmodel has been advanced to explain the molecular basis of early membrane modification by virus particles (Carrasco, 1994). The viral molecules involved are components of virions:glycoproteins when enveloped particles are analyzed or, still unidentified, domains of thestructural proteins in the case of naked viruses. Attachment of the particle to the cell surfacereceptor does not alter membrane permeability by itself. Inhibitors that hamper virus decap-sidation, still allowing virus attachment to the cell surface, block early membrane permeabi-lization (Almela et al., 1991).
At late times of infection, when there is active translation of late viral mRNAs, the
plasma membrane becomes permeable to small molecules and ions (Carrasco, 1978)(Figure 6.1). Different viral molecules may be responsible for this late enhancement of mem-brane permeability, including viroporins (Gonzalez and Carrasco, 2003), glycoproteins, andeven proteases (Chang et al., 1999; Blanco et al., 2003). This chapter is devoted to reviewingsome characteristics of membrane permeabilization by viral proteins. In addition, the method-ology used to assay enhanced permeability in animal cells is described. Finally, the design ofselective viral inhibitors based on the modification of cellular membranes during virus entryor at late times of infection is also discussed. María Eugenia González
Unidad de Expresión Viral, Centro Nacional de Microbiologia, Instituto de Salud
Carlos III, Carretera de Majadahonda-Pozuelo, Km 2, Majadahonda 28220, Madrid, Spain. Luis Carrasco
Centro de Biología Molecular Severo Ochoa, Facultad de Ciencias, Universidad Autónoma,
Viral Membrane Proteins: Structure, Function, and Drug Design, edited by Wolfgang Fischer. Kluwer Academic / Plenum Publishers, New York, 2004. María Eugenia González and Luis Carrasco Figure 6.1.
(A) Schematic representation of early membrane permeability. The different steps of virus attachment,
entry, and fusion events are shown. The protein toxin alpha-sarcin co-enters endosomes in conjunction with animalvirus particles. The fusion of the viral membrane with the endosome membrane induces the release of alpha-sarcinto the cytoplasm. (B) Permeabilization of the plasma membrane at late times of infection. The figure depicts the entryof the low molecular weight translation inhibitor hygromycin B through pores created at the plasma membrane. Alpha-sarcin is unable to pass through these pores. 2. Measuring Alterations in Membrane Permeability 2.1. The Hygromycin B Test
A number of hydrophilic molecules, including some antibiotics, poorly permeate
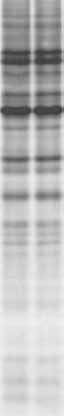
through cellular membranes (Contreras et al., 1978; Lacal et al., 1980). This is the case ofhygromycin B, anthelmycin, blasticidin S, destomycin A, gougerotin, and edein complex. The aminoglycoside antibiotic hygromycin B (MW 527) is produced by Streptomyceshygroscopicus. This hydrophilic molecule is an efficient inhibitor of protein synthesis in cell-free systems but interferes very poorly with translation in intact cells. However, themodification of the plasma membrane by viruses or by other means leads to a rapid blockadeof translation (Carrasco, 1995). Concentrations of the antibiotic ranging from about0.1–1 mM are added to the culture medium and protein synthesis is estimated by incubationwith radioactive methionine for 1 hr (see Figure 6.2) (Gonzalez and Carrasco, 2001). Inaddition to its simplicity, the hygromycin B test has a number of advantages for assayingchanges in membrane permeability. One is its great sensitivity, and another is that thistest measures membrane modifications only in cells that are metabolically active. Moreover,in cultures where some cells are uninfected, hygromycin B would only enter virus-infectedcells that are synthesizing proteins. The hygromycin B test has been applied withsuccess to prokaryotic (Lama and Carrasco, 1992) and eukaryotic cells, including yeast(Barco and Carrasco, 1995, 1998) and mammalian cells (Gatti et al., 1998; Gonzalez andCarrasco, 2001). Viral Proteins that Enhance Membrane Permeability Figure 6.2.
Entry of hygromycin B into BHK cells promoted by the expression of HIV-1 Vpu. Control BHK cells
(left), cells infected with Sindbis virus lacking the 6K gene (SV(-6K)) (center), or with (SV(-6K)) containing theHIV-1 vpu gene (SV(-6K)ϩVpu) (right) were treated or not with 0.5 mM hygromycin B for 10 min and proteinsynthesis was assayed by extended incubation with [35S] Met/Cys for 1 hr. The labeled proteins were analyzed byautoradiography after SDS PAGE. For additional details see Gonzalez and Carrasco (2001). 2.2. Entry of Macromolecules into Virus-Infected Cells
Alpha-sarcin is a protein of 150 amino acid residues, which is produced by Aspergillusgiganteus (Oka et al., 1990). This protein inhibits translation by modifying ribosomes in anenzymatic manner. Thus, a molecule of alpha-sarcin is able to inactivate a great number ofribosomes by hydrolysis of the A4324-G4325 phosphodiester bond in the 28S rRNA (Chanet al., 1983). This toxin does not enter mammalian cells because it does not attach to the cellsurface and is therefore unable to cross the plasma membrane. However, alpha-sarcin effi-ciently interferes with protein synthesis in cell-free systems or in cells where the permeabil-ity barrier has been destroyed (Fernandez-Puentes and Carrasco, 1980). Alpha-sarcinco-enters cells in conjunction with virus particles, and is liberated to the cytoplasm (Otero andCarrasco, 1987; Liprandi et al., 1997). In this manner, this toxin irreversibly blocks transla-tion several minutes after virus entry. The molecular basis of the co-entry of macromoleculeswith virus particles has been analyzed in detail elsewhere (Carrasco, 1994, 1995). Apart fromalpha-sarcin, a number of proteins that interfere with translation, many of them of plant origin, have been described (Fernandez-Puentes and Carrasco, 1980; Lee et al., 1990). Therelease of all these toxins into cells is enhanced by virus particles. Not only proteins, but also
María Eugenia González and Luis Carrasco
other macromolecules, including nucleic acids efficiently co-enter with virus particles(Cotten et al., 1992). However, none of these macromolecules passes into cells at late timesof infection. 2.3. Other Assays to Test the Entry or Exit of Molecules from Virus-Infected Cells
Apart from the use of translation inhibitors that do not easily permeate into intact cells,
a number of assays can be employed to assess modifications in membrane permeability. Amongst these assays, we can list the following.
2.3.1. Entry or Exit of Radioactive Molecules
Cells are preloaded with radioactive uridine and the exit of nucleotides can be moni-
tored after induction of viroporin expression (Gonzalez and Carrasco, 1998). Unlike withmost amino acids, the pool of uridine nucleotides is abundant in the cell interior, thus providing a convenient and sensitive assay for monitoring the exit of molecules from cells. Other tests use radioactive glucose derivatives (e.g., 2-deoxyglucose), which cannot bemetabolized and accumulate in cells. The analysis of the release of radioactive compoundsthat are not actively transported into cells leads to the failure to measure enhanced membranepermeability.
Entry of o-nitrophenyl--D-galactopyranoside (ONPG) into the bacterial cells can be
determined very simply. This -galactosidase substrate can be incubated with bacterial cellsand production of the resulting compound can be followed by determining the absorbance at 420 nm (Lama and Carrasco, 1992). There are a number of non-vital dyes employed to characterize cell mortality. It should be noted that the entry of these compounds, in fact,determines a modification in cell membranes, which in some cases does not directly correlatewith cell death. Trypan blue is a dye widely used for monitoring enhanced membrane perme-ability. However, this assay is not very sensitive. In addition, trypan blue staining does not dis-criminate between metabolically active or dead cells, as the hygromycin B tests does. Anotherassay employs the polyamine neurobiotin that needs specific connexin channels to enter mam-malian cells (Elfgang et al., 1995). Permeabilization of cell membrane increases uptake of thiscationic molecule. Internalized neurobiotin can be detected in paraformaldehyde-fixed cells,by fluorescence microscopy, using fluorescein isothiocyanate-conjugated streptavidin(Gonzalez and Carrasco, 1998).
Propidium iodide (PI) is a DNA-intercalating compound that does not enter intact cells.
However, those cells that exhibit increased membrane permeability are able to take up PI,which can be assayed by cell fluorometric analysis (Arroyo et al., 1995).
2.3.4. Release of Cellular Enzymes to the Culture Medium
The appearance in the culture medium of cellular enzymes is a clear indicator of
cell mortality. This is the case for lactic dehydrogenase and bacterial -galactosidase, present
Viral Proteins that Enhance Membrane Permeability
outside the cells (Sanderson et al., 1996). Commercial kits to measure this enzymatic activity are available. The release of cellular proteins to the medium occurs at very late timesof viral infection, when cells have already died. This alteration takes place at much later times after hygromycin B entry can be detected (Blanco et al., 2003). 3. Viral Proteins that Modify Permeability 3.1. Viroporins
Viroporins are small proteins encoded by viruses that contain a stretch of hydrophobic
amino acids (Gonzalez and Carrasco, 2003). Typically, viroporins are comprised of some60–120 amino acids. The hydrophobic domain is able to form an amphipathic ␣-helix. Theinsertion of these proteins into membranes followed by their oligomerization creates ahydrophilic pore. The architecture of this channel is such that the hydrophobic amino acidresidues face the phospholipid bilayer while the hydrophilic residues form part of the pore. In addition to this domain, there are other features of viroporin structure, including a secondhydrophobic region in some viroporins that also interacts with membranes. This second interaction may further disturb the organization of the lipid bilayer. These proteins may alsocontain a stretch of basic amino acids that acts in a detergent-like fashion. All these structuralfeatures contribute to membrane destabilization. More recently, another domain has beendescribed in some glycoproteins and also viroporins that has the capacity to interact withmembranes. This domain is rich in aromatic amino acids and is usually inserted at the inter-face of the phospholipid bilayer (Suarez et al., 2000; Sanz et al., 2003). This type of interac-tion also leads to membrane destabilization, further enhancing membrane permeability.
A number of viroporins from different viruses that infect eukaryotic cells have been
reported. This group of proteins includes picornavirus 2B and 3A, alphavirus 6K, retrovirusVpu, paramixovirus SH, orthomixovirus M2, reovirus p10, flavivirus p7, phycodnavirus Kcv,coronavirus E, and rhabdovirus alpha 10p. A recent review devoted to viroporins discussesthe structure and function of a number of proteins of this group (Gonzalez and Carrasco,2003), and so the details of each particular viroporin will not be reviewed in this chapter.
The main activity of viroporins is to create pores at biological membranes to permit the
passage of ions and small molecules. The cloning and individual expression of viroporingenes has allowed their effects in bacterial and animal cells to be analyzed. Thus, the expres-sion of this type of viral gene enhances the permeation of ions and several hydrophilic mole-cules in or out of cells (Carrasco, 1995). In addition, the purified viroporin molecules openpores in model membranes, providing a system that is amenable to biophysical analysis(Fischer and Sansom, 2002). The pore size created by viroporins allows the diffusion of different molecules with a molecular weight below about 1,000 Da.
The main step affected in animal viruses containing a deleted viroporin gene is the
assembly and exit of virions from the infected cells (Klimkait et al., 1990; Liljestrom et al.,1991; Loewy et al., 1995; Betakova et al., 2000; Watanabe et al., 2001; Kuo and Masters,2003). These genes are not essential for virus replication in culture cells, but the plaque sizeis much smaller in viroporin-defective viruses. Notably, virus entry and gene expression inviroporin-deleted viruses occur as in their wild-type counterparts. An aspect of viroporinfunction at the molecular level that is still not understood is the link between pore activity andvirus budding. María Eugenia González and Luis Carrasco 3.2. Viral Glycoproteins that Modify Membrane Permeability
In addition to small hydrophobic viral proteins, there are other virus products that
promote membrane permeabilization. This occurs with a number of virus glycoproteins (GP)that are known to increase cell membrane permeability, such as the human immunodeficiencyvirus gp41 (Chernomordik et al., 1994; Arroyo et al., 1995), the Ebola virus GP (Yang et al.,2000), the cytomegalovirus US9 protein (Maidji et al., 1996), the Vaccinia virus A38L pro-tein (Sanderson et al., 1996), rotavirus VP7 and NS4 proteins (Charpilienne et al., 1997;Newton et al., 1997), the hepatitis C virus E1 protein (Ciccaglione et al., 1998), and thealphavirus E1 protein (Nyfeler et al., 2001; Wengler et al., 2003).
The architecture of some viral glycoproteins is such that upon oligomerization, the
transmembrane (TM) domains may form a physical pore. In principle, two different regionsof a viral fusion glycoprotein could form pores. One such region contains the fusion peptidethat would create a pore in the cell membranes upon insertion (Skehel and Wiley, 1998), whilethe TM domain would form a pore in the virion membrane (Wild et al., 1994). Moreover,sequences adjacent to the TM region could have motifs designed to destabilize membranestructure (Suarez et al., 2000). Entry of enveloped animal viruses leads to early membranepermeabilization, which is mediated by the formation of the two pores (fusion and TM)formed by viral fusion glycoproteins. This early permeabilization induced during the entry ofvirions requires conformational changes of the fusion glycoproteins. By contrast, after virusreplication, newly synthesized glycoproteins may affect membrane permeability when theyreach the plasma membrane (Figure 6.3). This modification is achieved only by the TMdomain, while the fusion peptide does not participate in this late modification. In viruses thatlack the typical viroporin, its function could be replaced by these pore-forming glycoproteins,while for other viruses viroporin activity may be redundant (Bour and Strebel, 1996). In thelatter case, pore formation may be generated by viral glycoproteins and viroporins(Figure 6.3). We would like to propose the possibility that pore-forming glycoproteins play a key role mainly during virus entry and, in some cases, also during virus budding, while viroporins come into action when viruses need to exit the cell.
Early membrane permeabilization is always carried out by a virion component. In the
case of enveloped viruses, this early event is executed by a structural glycoprotein, which iscoupled to the fusion process. An understanding of fusion at the molecular level also requiresan explanation of the phenomenon of early membrane permeabilization. We have advancedthe idea that viral glycoproteins involved in membrane fusion participate in the dissipation of the chemiosmotic gradient, thus providing the energy to push the nucleocapsid and neighboring macromolecules to the cell interior (Carrasco, 1994; Irurzun et al., 1997). Fusionglycoproteins do not simply serve to bridge the cellular and the viral membrane, but insteadare designed to open pores in both membranes. This pore-opening activity may be necessaryto lower membrane potential and to dissipate ionic gradients. Several chapters of this bookare devoted to the detailed description of the structure and function of these glycoproteins, sowe will focus our attention on viral glycoproteins that permeabilize membranes when indi-vidually expressed in cells. These membrane active proteins may exhibit this activity later onin the virus life cycle.
Rotavirus infection provokes a number of alterations in cellular membranes during
infection (del Castillo et al., 1991). Amongst these alterations, there is an increase in the
Viral Proteins that Enhance Membrane Permeability Figure 6.3.
Participation of pore formation by viral glycoproteins and viroporins in membrane permeability.
Early membrane permeabilization is coupled to the fusion activity of the corresponding viral glycoprotein. Thisfusion glycoprotein may create two pores. One is located at the target cell membrane and the other is formed by the TM domain. Late membrane permeabilization may be carried out by viroporins or by the TM domains of viralglycoproteins.
concentration of cytoplasmic calcium (Michelangeli et al., 1991). Several rotavirus proteinsexhibit membrane-destabilizing activity. The enterotoxin NSP4 induces alterations in mem-brane permeability (Tian et al., 1994). The individual expression of the non-structural glyco-protein NSP4 has the ability to increase the concentration of cytoplasmic calcium. Thisincrease may be mediated by activation of phospholipase C activity (Dong et al., 1997).
Rotavirus particles induce the co-entry of protein toxins into cells (Cuadras et al.,
1997). At least two structural components possess the ability to permeabilize cells, includingVP5 protein and VP7 glycoprotein (Charpilienne et al., 1997; Irurzun et al., 1997).
Infection of lymphocytic human cells by HIV-1 enhances membrane permeability
to ions and several compounds (Voss et al., 1996; Gatti et al., 1998). There are at least threedifferent HIV-encoded proteins responsible for these alterations: Vpu protein, the retroviralprotease, and the fusion glycoprotein gp41. Apart from the fusion peptide, there are tworegions of gp41 that exhibit membrane permeability; one is located at the carboxy terminus(Arroyo et al., 1995; Comardelle et al., 1997) and another corresponds to the membrane-spanning domain (Arroyo et al., 1995). The C-terminus of gp41 includes two 20–30 residues,which may form cationic amphipathic ␣-helices, designated as lentivirus lytic peptides 1 and 2(LLP-1 and LLP-2). Synthetic LLP-1 peptide forms pores in planar phospholipid bilayers
María Eugenia González and Luis Carrasco
(Chernomordik et al., 1994), permeabilizes HIV-1 virions to deoxyribonucleoside triphos-phates (Zhang et al., 1996), and induces alterations in ion permeability of Xenopus oocytes(Comardelle et al., 1997).
Inducible expression of the hepatitis C virus E1 glycoprotein increases membrane
permeability in bacterial cells. The ability of E1 to modify membrane permeability has beenmapped to the carboxy terminus of the protein (Ciccaglione et al., 1998, 2001). Similar permeabilization was found with Escherichia coli cells that synthesize Semliki forest virus E1glycoprotein after exposure to low pH (Nyfeler et al., 2001). Finally, overexpression of Vacciniavirus A38L glycoprotein produces changes in the morphology, permeability, and adhesion ofmammalian cells. The potential capacity of A38L protein to form pores at the plasma membranepromotes the entry of calcium ions and PI and the release of lactic dehydrogenase into theculture medium (Sanderson et al., 1996). 4. Membrane Permeabilization and Drug Design 4.1. Antibiotics and Toxins that Selectively Enter Virus-Infected Cells
Different approaches have been envisaged for the design of compounds that interfere
with virus replication based on modifications in membrane permeability. One such approachmakes use of inhibitors of cellular or viral functions that do not permeate easily into intactanimal cells. Notably, these agents selectively enter into virus-infected cells (Carrasco, 1978;Benedetto et al., 1980). Most of the inhibitors used thus far interfere with protein synthesis,although compounds that affect other functions could also be employed. Entry of these agentsleads to the inhibition of translation specifically in virus-infected cells, leading to a profoundinhibition of virus growth (Contreras et al., 1978; Carrasco, 1995; Gatti et al., 1998). Although this approach discriminates well between uninfected or virus-infected cells in cul-ture, the high toxicity of the agents thus far assayed has hampered its use in whole animals. Perhaps future searches for less toxic compounds would make this approach amenable toapplication in therapy. In fact, some of the plant toxins that co-enter with virus particles havebeen described as being antiviral agents (Lee et al., 1990). Even compounds such ashygromycin B, which has been used in the veterinary field as an antibacterial agent, couldalso be used as an antiviral compound for rotavirus infections (Liprandi et al., 1997). 4.2. Viroporin Inhibitors
The paradigm of an inhibitor of a viral ion channel is amantadine (Hay, 1992)
(Figure 6.4). This compound has been used as an anti-influenza agent in humans (De Clercq,2001). The target of amantadine is the influenza-encoded protein M2. Residues 27, 30, 31,and 34 of M2 determine the amantadine sensitivity of this ion channel. A drawback ofamantadine is the high doses necessary to affect influenza. The search for more effectivecompounds may provide a more efficacious treatment for this illness.
Compounds that interfere with the functioning of other viroporins have also been
described. This is the case of amiloride derivatives that block HIV-1 Vpu activity (Ewart et al.,
Viral Proteins that Enhance Membrane Permeability Figure 6.4.
Chemical structures of several inhibitors of viroporin activity.
2002). In this manner, the production of infectious HIV-1 is reduced in the presence of thisagent. Recently, long alkyl-chain iminosugar derivatives have been found to interfere with thefunction of the hepatitis C virus p7 protein as an ion channel (Griffin et al., 2003; Pavlovicet al., 2003). These compounds exhibited antiviral activity with bovine viral diarrhea virus,which is closely related to the hepatitis C virus (Durantel et al., 2001). 4.3. Antiviral Agents that Interfere with Viral Glycoproteins
Much effort has been concentrated recently on the development of antiviral agents that
inhibit the fusion step of HIV. Binding of HIV gp120 to the CD4 receptor is followed byfurther interaction of this viral glycoprotein with the coreceptor molecules CXCR4 and CCR5. After this initial interaction, the conformation of the ectodomain of the TM glycoprotein gp41is profoundly modified. Exposure of the fusion peptide at the amino terminus of gp41 triggersits insertion into the target cellular membrane, leading to the fusion of the viral and the cellularplasma membranes. All these steps have been used as targets for anti-HIV therapy (Cooley andLewin, 2003). As regards the fusion step, a variety of peptide mimetic inhibitors havebeen developed. The pioneering work on peptide T20 has demonstrated that this compoundis a potent inhibitor of gp41-induced membrane fusion. T20 exhibits antiviral activity in HIV-infected patients. The detailed mechanism of action of T20 at the molecular level isknown. This peptide is homologous with 36 amino acids within the C-terminal heptad repeatregion (HR2) of HIV-1 gp41. T20 competitively binds to HR1 and interferes with the forma-tion of the six helix HR1–HR2 bundle complex necessary for membrane fusion. At presentthere are a great number of peptides that interfere with binding of gp120 or with gp41-inducedmembrane fusion and that have been tested for their anti-HIV activity and clinical efficacy. Inthis regard, T1249 is one of the second generation of HR-2 peptide mimetic inhibitors thatconsists of 39 amino acids. PRO 542 is a soluble CD4 receptor (CD4-IgG2) that binds toand neutralizes gp120 before virus binding occurs. SCH-C is an oxime–piperidine compoundthat is a coreceptor antagonist. This small molecule acts as an inhibitor of CCR5. MD3100 is
María Eugenia González and Luis Carrasco
a non-peptidic, low molecular weight bicyclam compound that prevents interactions betweenCXCR4 and gp120, blocking signal transduction from CXCR4. Future research in this fieldwill provide us with additional antiviral compounds to add to the anti-HIV armory. Acknowledgments
This work was supported by grants from the Fundación para la Investigación y
Prevención del SIDA en España (24291), Instituto de Salud Carlos III (01/0042), and theDGICYT PM99-0002. The authors also acknowledge the Fundación Ramón Areces for aninstitutional grant awarded to the Centro de Biología Molecular “Severo Ochoa.”
References
Almela, M.J., Gonzalez, M.E., and Carrasco, L. (1991). Inhibitors of poliovirus uncoating efficiently block the early
membrane permeabilization induced by virus particles. J. Virol. 65, 2572–2577.
Arroyo, J., Boceta, M., Gonzalez, M.E., Michel, M., and Carrasco, L. (1995). Membrane permeabilization by
different regions of the human immunodeficiency virus type 1 transmembrane glycoprotein gp41. J. Virol. 69, 4095–4102.
Barco, A. and Carrasco, L. (1995). A human virus protein, poliovirus protein 2BC, induces membrane proliferation
and blocks the exocytic pathway in the yeast Saccharomyces cerevisiae. EMBO J. 14, 3349–3364.
Barco, A. and Carrasco, L. (1998). Identification of regions of poliovirus 2BC protein that are involved in cytotoxi-
city. J. Virol. 72, 3560–3570.
Benedetto, A., Rossi, G.B., Amici, C., Belardelli, F., Cioe, L., Carruba, G. et al. (1980). Inhibition of animal virus
production by means of translation inhibitors unable to penetrate normal cells. Virology 106, 123–132.
Betakova, T., Wolffe, E.J., and Moss, B. (2000). The vaccinia virus A14.5L gene encodes a hydrophobic
53-amino-acid virion membrane protein that enhances virulence in mice and is conserved among vertebrate poxviruses. J. Virol. 74, 4085–4092.
Blanco, R., Carrasco, L., and Ventoso, I. (2003). Cell killing by HIV-1 protease. J. Biol. Chem. 278, 1086–1093. Bour, S. and Strebel, K. (1996). The human immunodeficiency virus (HIV) type 2 envelope protein is a functional
complement to HIV type 1 Vpu that enhances particle release of heterologous retroviruses. J. Virol. 70, 8285–8300.
Carrasco, L. (1978). Membrane leakiness after viral infection and a new approach to the development of antiviral
agents. Nature 272, 694–699.
Carrasco, L. (1994). Entry of animal viruses and macromolecules into cells. FEBS Lett. 350, 151–154. Carrasco, L. (1995). Modification of membrane permeability by animal viruses. Adv. Virus Res. 45, 61–112. Chan, Y.L., Endo, Y., and Wool, I.G. (1983). The sequence of the nucleotides at the alpha-sarcin cleavage site in rat
28S ribosomal ribonucleic acid. J. Biol. Chem. 258, 12768–12770.
Chang, Y.S., Liao, C.L., Tsao, C.H., Chen, M.C., Liu, C.I., Chen, L.K. et al. (1999). Membrane permeabilization by
small hydrophobic nonstructural proteins of Japanese encephalitis virus. J. Virol. 73, 6257–6264.
Charpilienne, A., Abad, M.J., Michelangeli, F., Alvarado, F., Vasseur, M., Cohen, J. et al. (1997). Solubilized and
cleaved VP7, the outer glycoprotein of rotavirus, induces permeabilization of cell membrane vesicles. J. Gen. Virol. 78, 1367–1371.
Chernomordik, L., Chanturiya, A.N., Suss-Toby, E., Nora, E., and Zimmerberg, J. (1994). An amphipathic peptide
from the C-terminal region of the human immunodeficiency virus envelope glycoprotein causes pore formation in membranes. J. Virol. 68, 7115–7123.
Ciccaglione, A.R., Costantino, A., Marcantonio, C., Equestre, M., Geraci, A., and Rapicetta, M. (2001). Mutagenesis
of hepatitis C virus E1 protein affects its membrane-permeabilizing activity. J. Gen. Virol. 82, 2243–2250.
Ciccaglione, A.R., Marcantonio, C., Costantino, A., Equestre, M., Geraci, A., and Rapicetta, M. (1998). Hepatitis C
virus E1 protein induces modification of membrane permeability in E. coli cells. Virology 250, 1–8.
Comardelle, A.M., Norris, C.H., Plymale, D.R., Gatti, P.J., Choi, B., Fermin, C.D. et al. (1997). A synthetic peptide cor-
responding to the carboxy terminus of human immunodeficiency virus type 1 transmembrane glycoprotein induces alterations in the ionic permeability of Xenopus laevis oocytes. AIDS Res. Hum. Retroviruses 13, 1525–1532. Viral Proteins that Enhance Membrane Permeability
Contreras, A., Vazquez, D., and Carrasco, L. (1978). Inhibition, by selected antibiotics, of protein synthesis in cells
growing in tissue cultures. J. Antibiot. (Tokyo) 31, 598–602.
Cooley, L.A. and Lewin, S.R. (2003). HIV-1 cell entry and advances in viral entry inhibitor therapy. J. Clin. Virol.26, 121–132.
Cotten, M., Wagner, E., Zatloukal, K., Phillips, S., Curiel, D.T., and Birnstiel, M.L. (1992). High-efficiency receptor-
mediated delivery of small and large (48 kilobase) gene constructs using the endosome-disruption activity of defective or chemically inactivated adenovirus particles. Proc. Natl. Acad. Sci. USA 89, 6094–6098.
Cuadras, M.A., Arias, C.F., and Lopez, S. (1997). Rotaviruses induce an early membrane permeabilization of MA104
cells and do not require a low intracellular Caϩ concentration to initiate their replication cycle. J. Virol. 71,
De Clercq, E. (2001). Antiviral drugs: Current state of the art. J. Clin. Virol. 22, 73–89. del Castillo, J.R., Ludert, J.E., Sanchez, A., Ruiz, M.C., Michelangeli, F., and Liprandi, F. (1991). Rotavirus infection
alters Naϩ and Kϩ homeostasis in MA-104 cells. J. Gen. Virol. 72, 541–547.
Dong, Y., Zeng, C.Q., Ball, J.M., Estes, M.K., and Morris, A.P. (1997). The rotavirus enterotoxin NSP4 mobilizes intra-
cellular calcium in human intestinal cells by stimulating phospholipase C-mediated inositol 1,4,5-trisphosphate production. Proc. Natl. Acad. Sci. USA 94, 3960–3965.
Durantel, D., Branza-Nichita, N., Carrouee-Durantel, S., Butters, T.D., Dwek, R.A., and Zitzmann, N. (2001). Study
of the mechanism of antiviral action of iminosugar derivatives against bovine viral diarrhea virus. J. Virol. 75, 8987–8998.
Elfgang, C., Eckert, R., Lichtenberg-Frate, H., Butterweck, A., Traub, O., Klein, R.A. et al. (1995). Specific perme-
ability and selective formation of gap junction channels in connexin-transfected HeLa cells. J. Cell Biol. 129, 805–817.
Ewart, G.D., Mills, K., Cox, G.B., and Gage, P.W. (2002). Amiloride derivatives block ion channel activity and
enhancement of virus-like particle budding caused by HIV-1 protein Vpu. Eur. Biophys. J. 31, 26–35.
Fernandez-Puentes, C. and Carrasco, L. (1980). Viral infection permeabilizes mammalian cells to protein toxins. Cell20, 769–775.
Fischer, W.B. and Sansom, M.S. (2002). Viral ion channels: Sructure and function. Biochim. Biophys. Acta 1561,
Gatti, P.J., Choi, B., Haislip, A.M., Fermin, C.D., and Garry, R.F. (1998). Inhibition of HIV type 1 production by
hygromycin B. AIDS Res. Hum. Retroviruses 14, 885–892.
Gonzalez, M.E. and Carrasco, L. (1998). The human immunodeficiency virus type 1 Vpu protein enhances
membrane permeability. Biochemistry 37, 13710–13719.
Gonzalez, M.E. and Carrasco, L. (2001). Human immunodeficiency virus type 1 VPU protein affects Sindbis virus
glycoprotein processing and enhances membrane permeabilization. Virology 279, 201–209.
Gonzalez, M.E. and Carrasco, L. (2003). Viroporins. FEBS Lett. 552, 28–34. Griffin, S.D., Beales, L.P., Clarke, D.S., Worsfold, O., Evans, S.D., Jaeger, J. et al. (2003). The p7 protein of
hepatitis C virus forms an ion channel that is blocked by the antiviral drug, Amantadine. FEBS Lett. 535, 34–38.
Hay, A.J. (1992). The action of adamantanamines against influenza A viruses: Inhibition of the M2 ion channel
protein. Semin. Virol. 3, 21–30.
Irurzun, A., Nieva, J.L., and Carrasco, L. (1997). Entry of Semliki forest virus into cells: Effects of concanamycin A
and nigericin on viral membrane fusion and infection. Virology 227, 488–492.
Klimkait, T., Strebel, K., Hoggan, M.D., Martin, M.A., and Orenstein, J.M. (1990). The human immunodeficiency
virus type 1-specific protein vpu is required for efficient virus maturation and release. J. Virol. 64, 621–629.
Kuo, L. and Masters, P.S. (2003). The small envelope protein E is not essential for murine coronavirus replication. J. Virol. 77, 4597–4608.
Lacal, J.C., Vazquez, J.M., Fernandez-Sousa, D., and Carrasco, L. (1980). Antibiotics that specifically block transla-
tion in virus-infected cells. J. Antibiot. (Tokyo) 33, 441–446.
Lama, J. and Carrasco, L. (1992). Expression of poliovirus nonstructural proteins in Escherichia coli cells.
Modification of membrane permeability induced by 2B and 3A. J. Biol. Chem. 267, 15932–15937.
Lee, T., Crowell, M., Shearer, M.H., Aron, G.M., and Irvin, J.D. (1990). Poliovirus-mediated entry of pokeweed
antiviral protein. Antimicrob. Agents Chemother. 34, 2034–2037.
Liljestrom, P., Lusa, S., Huylebroeck, D., and Garoff, H. (1991). In vitro mutagenesis of a full-length cDNA clone
of Semliki Forest virus: The small 6,000-molecular-weight membrane protein modulates virus release. J. Virol. 65, 4107–4113.
Liprandi, F., Moros, Z., Gerder, M., Ludert, J.E., Pujol, F.H., Ruiz, M.C. et al. (1997). Productive penetration of
rotavirus in cultured cells induces co-entry of the translation inhibitor alpha-sarcin. Virology 237, 430–438. María Eugenia González and Luis Carrasco
Loewy, A., Smyth, J., von Bonsdorff, C.H., Liljestrom, P., and Schlesinger, M.J. (1995). The 6-kilodalton membrane
protein of Semliki Forest virus is involved in the budding process. J. Virol. 69, 469–475.
Maidji, E., Tugizov, S., Jones, T., Zheng, Z., and Pereira, L. (1996). Accessory human cytomegalovirus glycoprotein
US9 in the unique short component of the viral genome promotes cell-to-cell transmission of virus in polarized epithelial cells. J. Virol. 70, 8402–8410.
Michelangeli, F., Ruiz, M.C., del Castillo, J.R., Ludert, J.E., and Liprandi, F. (1991). Effect of rotavirus infection on
intracellular calcium homeostasis in cultured cells. Virology 181, 520–527.
Newton, K., Meyer, J.C., Bellamy, A.R., and Taylor, J.A. (1997). Rotavirus nonstructural glycoprotein NSP4 alters
plasma membrane permeability in mammalian cells. J. Virol. 71, 9458–9465.
Nyfeler, S., Senn, K., and Kempf, C. (2001). Expression of Semliki Forest virus E1 protein in Escherichia coli. Low
pH-induced pore formation. J. Biol. Chem. 276, 15453–15457.
Oka, T., Natori, Y., Tanaka, S., Tsurugi, K., and Endo, Y. (1990). Complete nucleotide sequence of cDNA for the
cytotoxin alpha sarcin. Nucleic Acids Res. 18, 1897.
Otero, M.J. and Carrasco, L. (1987). Proteins are cointernalized with virion particles during early infection. Virology160, 75–80.
Pavlovic, D., Neville, D.C., Argaud, O., Blumberg, B., Dwek, R.A., Fischer, W.B. et al. (2003). The hepatitis C virus
p7 protein forms an ion channel that is inhibited by long-alkyl-chain iminosugar derivatives. Proc. Natl. Acad. Sci. USA 100, 6104–6108.
Sanderson, C.M., Parkinson, J.E., Hollinshead, M., and Smith, G.L. (1996). Overexpression of the vaccinia virus
A38L integral membrane protein promotes Caϩ influx into infected cells. J. Virol. 70, 905–914.
Sanz, M.A., Madan, V., Carrasco, L., and Madan, J.L. (2003). Interfacial domains in Sindbis virus 6K protein.
Detection and functional characterization. J. Biol. Chem. 278, 2051–2057.
Skehel, J.J. and Wiley, D.C. (1998). Coiled coils in both intracellular vesicle and viral membrane fusion. Cell 95,
Suarez, T., Gallaher, W.R., Agirre, A., Goni, F.M., and Nieva, J.L. (2000). Membrane interface-interacting sequences
within the ectodomain of the human immunodeficiency virus type 1 envelope glycoprotein: Putative role during viral fusion. J Virol. 74, 8038–8047.
Tian, P., Hu, Y., Schilling, W.P., Lindsay, D.A., Eiden, J., and Estes, M.K. (1994). The nonstructural glycoprotein of
rotavirus affects intracellular calcium levels. J. Virol. 68, 251–257.
Voss, T.G., Fermin, C.D., Levy, J.A., Vigh, S., Choi, B., and Garry, R.F. (1996). Alteration of intracellular potassium
and sodium concentrations correlates with induction of cytopathic effects by human immunodeficiency virus. J. Virol. 70, 5447–5454.
Watanabe, T., Watanabe, S., Ito, H., Kida, H., and Kawaoka, Y. (2001). Influenza A virus can undergo multiple cycles
of replication without M2 ion channel activity. J. Virol. 75, 5656–5662.
Wengler, G., Koschinski, A., Wengler, G., and Dreyer, F. (2003). Entry of alphaviruses at the plasma membrane
converts the viral surface proteins into an ion-permeable pore that can be detected by electrophysiological analyses of whole-cell membrane currents. J. Gen. Virol. 84, 173–181.
Wild, C.T., Shugars, D.C., Greenwell, T.K., McDanal, C.B., and Matthews, T.J. (1994). Peptides corresponding to a
predictive alpha-helical domain of human immunodeficiency virus type 1 gp41 are potent inhibitors of virus infection. Proc. Natl. Acad. Sci. USA 91, 9770–9774.
Yang, Z.Y., Duckers, H.J., Sullivan, N.J., Sanchez, A., Nabel, E.G., and Nabel, G.J. (2000). Identification of the
Ebola virus glycoprotein as the main viral determinant of vascular cell cytotoxicity and injury. Nat. Med. 6, 886–889.
Zhang, H., Dornadula, G., Alur, P., Laughlin, M.A., and Pomerantz, R.J. (1996). Amphipathic domains in the
C terminus of the TM protein (gp41) permeabilize HIV-1 virions: A molecular mechanism underlying natural endogenous reverse transcription. Proc. Natl. Acad. Sci. USA 93, 12519–12524.
Analgesics for “Go Home” post Spay-Neuter The following agents can be used for “go home” analgesics for spay and neuter patients : CANINE Analgesic agents for go home use : Realizing that an injection or oral administration of a separate analgesic may not be necessary depending on patient status, surgical technique, anesthesia/analgesic surgical protocol and perioperative drug use

 Viral Proteins that Enhance Membrane Permeability
Viral Proteins that Enhance Membrane Permeability